

Body
The development of recombinant DNA technologies and Agrobacterium-mediated transformation enabled expression of foreign genes in plants. Two main expression strategies have been utilized for expression of foreign gene products in plants: i) transient expression through either agroinfiltration or plant virus-based vector and ii) stable transgenic expression by insertion of foreign genes into the plant genome. Plant virus-based vector systems have been revealed to be more beneficial for relatively high level accumulation and rapid production of foreign proteins than regeneration of transgenic plant (Dawson and Folimonova, 2013; Gleba et al., 2007; Hefferon, 2012; Pogue et al., 2002; Shamekova et al., 2014). Moreover, these plant virus vectors can be used for application on other susceptible plants without additional genetic transformation of each host plants (Scholthof et al., 2002; Seaberg et al., 2012).
Plant virus-based vector systems can also be utilized to knock down expression of endogenous genes in plants. This virus-induced gene silencing (VIGS) technique has been widely used to elucidate gene function(s) in plants (Burch-Smith et al., 2004; Kurth et al., 2012; Muruganantham et al., 2009). Even though many VIGS vectors have been developed in recent years, host range of these vectors were mainly restricted in annual or herbaceous plants (Brigneti et al., 2004; Fofana et al., 2004; Holzberg et al., 2002; Kang et al., 2016; Kumagai et al., 1995; Liu et al., 2002). Here, we used grapevine Algerian latent virus (GALV) which can infects both perennial grape varieties and herbaceous Nicotiana benthamiana plants and tested its ability for VIGS vector.
GALV is a member of the genus Tombusvirus, family Tombusviridae and was first isolated in Italy from an Algerian vine (Gallitelli et al., 1989). GALV has subsequently been detected from water samples in agricultural areas, Gypsophila paniculata, nipplefruit, and Limonium sinuatum (Cannizzaro et al., 1990; Fuchs et al., 1994; Fujinaga et al., 2009; Kim et al., 2012; Koenig et al., 2004; Ohki et al., 2006; Yi et al., 1992). Grapevine plants infected by GALV show different symptoms such as chlorotic spots around the veins, mild vein clearing or mottling, malformations or curling of the leaf, and dark green blistering on the leaves in different genotypes (Lovato et al., 2014).
The grape is one of the earliest domesticated fruit crops and the world’s most valuable horticultural crop for the production of wine and table grapes (Myles et al., 2011). With the importance of grapevines, virus-based vectors could provide diverse opportunities and novel possibilities as biotechnological tools for plant improvement or disease protection. There have been only two virus-based vector systems for grapevine plants including grapevine virus A (GVA) and grapevine leafroll-associated virus-2 (GLRaV-2) (Haviv et al., 2006; Kurth et al., 2012; Muruganantham et al., 2009). Diverse virus-derived vector system is required for grape varieties because stability and effectiveness of virus-based vectors depends on both plant traits and virus vectors. In this study, we constructed a GAVL coat protein (CP)-replacement vector system and used to express the foreign gene and knock down the endogenous gene in the model plant N. benthamiana.
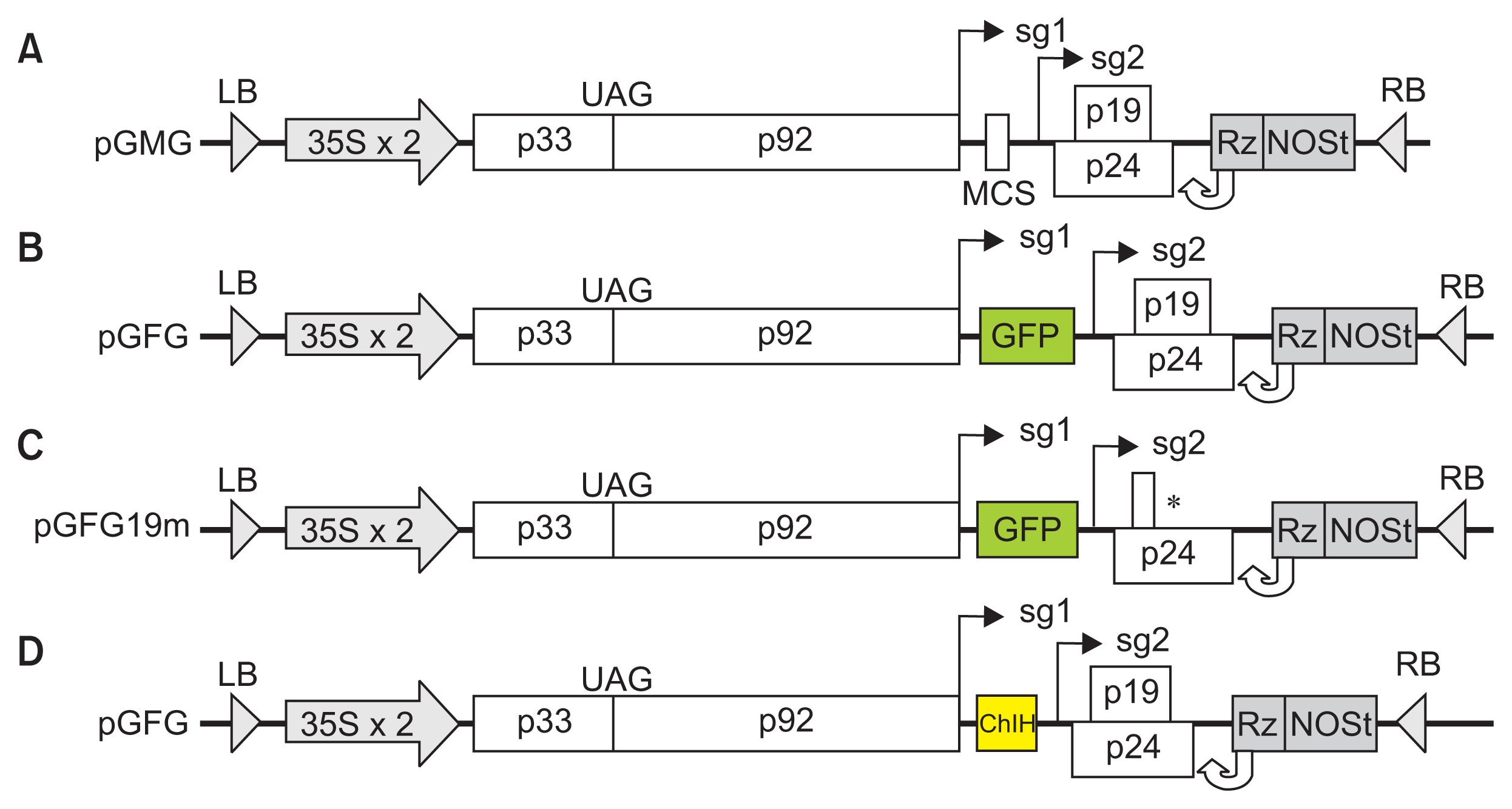
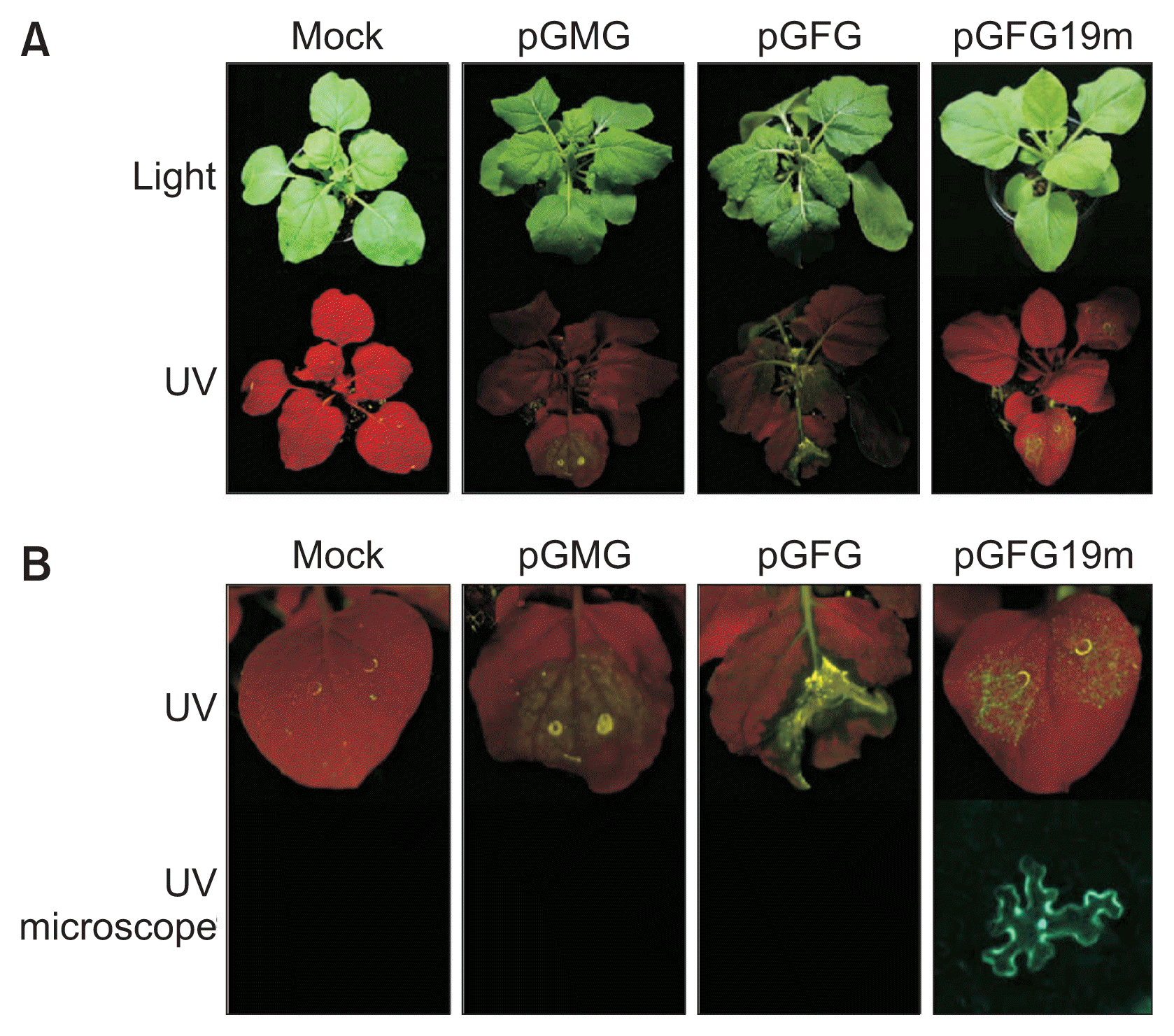
We previously constructed the CP deletion infectious clone of GALV, pGMG (Fig. 1) and confirmed systemic infection in N. benthamiana (Kim et al., 2012). To evaluate the GALV clone as an expression vector, a green fluorescence protein (GFP) sequence was used as a reporter gene which is expressed from the subgenomic promoter, and Agrobacterium-mediated gene delivery system was utilized for inoculation of N. benthamiana plants. A PCR product (GFP; about 700 bp) was amplified from pPZP-GFP (Powers et al., 2008) by using specific primers, GFP-5-MluI and GFP-3-MluI (Table 1). The PCR product was digested by MluI and inserted into pGMG resulting in pGFG (Fig. 1). Typical GALV systemic symptoms were observed from both pGMG and pGFG-inoculated plants as shown previously (Kim et al., 2012). However, severe necrosis was induced on the inoculated leaves of pGMG and pGFG at 7 days post-infiltration (dpi) and the green fluorescence was not detected on the infiltrated leaves (Fig. 2). In other tombusviruses, the p19 protein of tomato bushy stunt virus (TBSV) and cymbidium ringspot virus is a symptom determinant and responsible for eliciting the severe systemic necrosis on N. benthamiana (Dalmay et al., 1993; Scholthof et al., 1995). To avoid the necrotic cell death of infiltrated leaves, mutation of the p19 coding region, which results in the premature stop of p19, was introduced into pGFG, pGFG19m, as described previously (Fig. 1) (Kim et al., 2012). The amino acid, Ile (isoleucine), at position 157 from the start codon of p24 was changed to a stop-codon by changing ATC to ATT resulting in inactivation of p19 without disruption of the amino acid sequence of p24. Briefly, the region of p19/p24 was amplified by PCR using GALV-5-KpnI and GALV-3-KpnI primers (Table 1), and the PCR product was cloned into pGEM-T Easy vector (Promega, Madison, WI, USA). This plasmid was used for site-directed mutagenesis and PCR was performed with the primers, GALV-sense-157 and GALV-antisense-157 (Table 1). The amplified product was treated with DpnI restriction enzyme (Roche Applied Science, Mannheim, Germany) to remove the template plasmid before the bacterial transformation of the DpnI-treated PCR mixture. The mutated sequence was confirmed by sequencing. As we expected, the green fluorescence was observed when pGFG19m was introduced to leaves (Fig. 2). However, the expression level of GFP was substantially low in the pGFG19m-infiltrated leaves and the fluorescence was abolished at 10 dpi (Fig. 3). This result suggests that wild type p19 inhibits the VIGS and cause severe necrosis, and therefore the GFP transgene was silenced by VIGS in the absence of P19 expression. To determine whether the significant attenuation of GFP expression is associated with RNA silencing and unrelated silencing suppressor can restore the GFP expression, HC-Pro from soybean mosaic virus, was co-infiltrated with pGFG19m. The result showed that the GFP expression of pGFG19m in the presence of HC-Pro was significantly increased when compared to that of pGFG19m at both 6 and 10 dpi (Fig. 3), demonstrating that the attenuation of GFP expression in pGFG19m infection is caused by RNA silencing. To test the stability of the foreign gene, viruses in plant sap were passaged three times from plant to plant by mechanical sap-inoculation. Total RNAs were prepared from each individual plant and analyzed for stable GFP gene insertion in the viral genome by RT-PCR using primers spanning the p92 and p24 cistrons. A unique band with the predicted size was observed for viral GALV-GFP from plants inoculated with progeny viruses through serial passages (data not shown). Taken together, we developed two kinds of the GALV-based virus vectors, pGMG and pGMGm19, which could apply onto herbaceous or grapevine plants and pGMGm19 can be used for expression of a transgene in N. benthamiana expressing HC-Pro in trans.
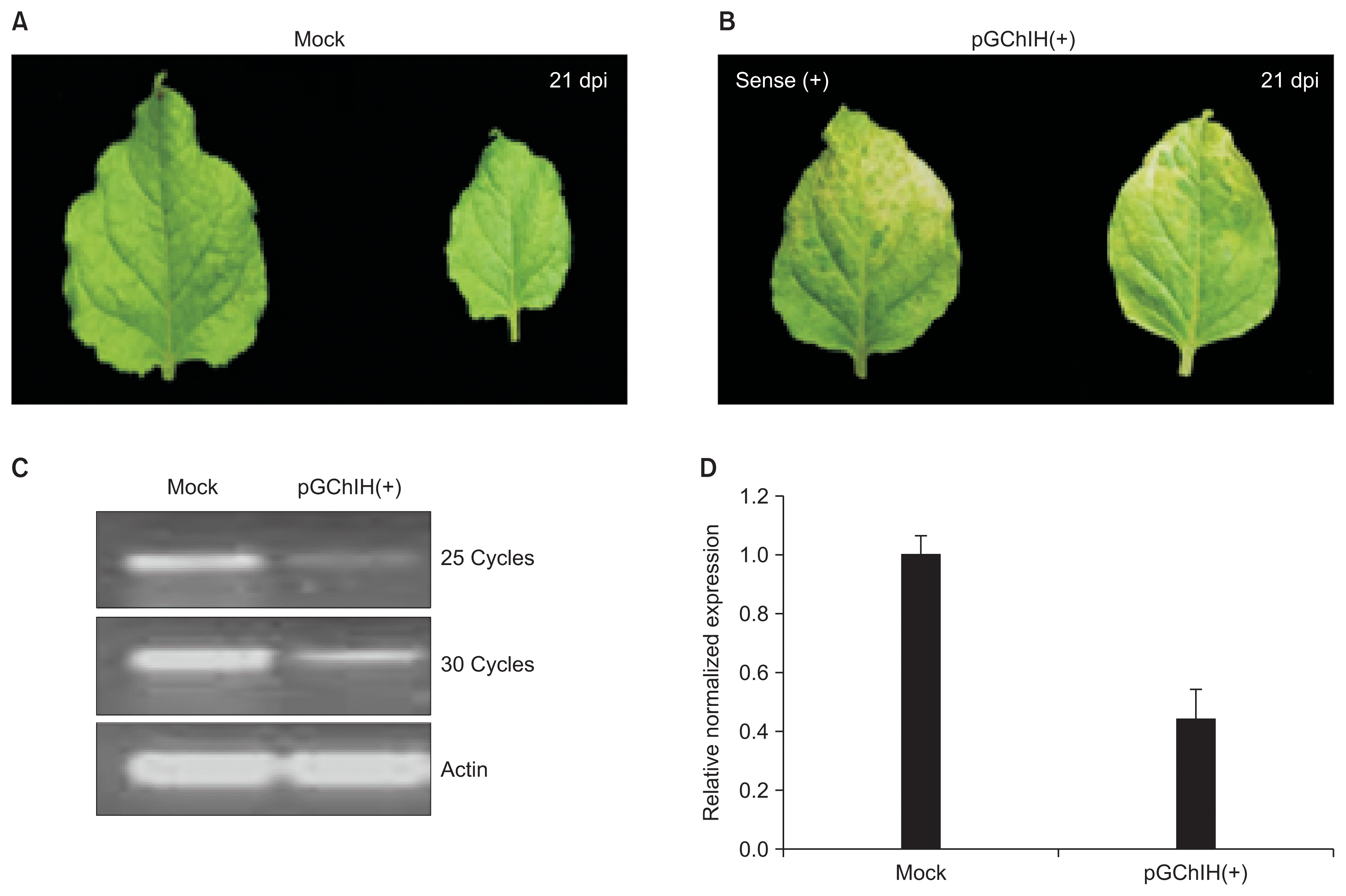
As the GALV-based vector effectively elicits RNA silencing in N. benthamiana, we decided to investigate the ability of GALV-based vector as a gene silencing vector. In other tombusvirus, CP replacement TBSV vectors were developed to knock down two endogenous genes, magnesium chelatase (ChlH) and phytoene desaturase (PDS) even though no silencing phenotype for PDS was observed (Pignatta et al., 2007). Total RNAs were extracted from N. benthamiana and cDNA was synthesized by using ChlH-3-MluI (Table 1). About 230 bp PCR fragment of ChlH was amplified with primers, ChlH-5-MluI and ChlH-3-MluI (Table 1), and the PCR product was inserted into the pGMG vector in the sense orientation resulting in pGChlH(+). The pGChlH(+) was inoculated in N. benthamiana and examined the silencing phenotype. At 12 to 14 dpi, chlorosis on newly leaves was observed (Fig. 4B). As expected, mRNA accumulation for ChlH gene was significantly lower in the leaves inoculated with pGChlH(+) than in that inoculated with mock (Fig. 4C, D). This result indicates that the ChlH gene was successfully silenced in N. benthamiana plants.
The development of a useful virus-based vector for expressing foreign genes and silencing endogenous genes is considered to be a valuable tool on the production of either useful genes or resistance genes and the study of gene function in the important crop. Until not so long ago, virus-based vectors are mainly applicable to annual plants. In grapevine as one of long-lived perennial crops, however, only two virus-derived vectors including GVA and GLRaV-2 were developed as mentioned above. GVA is a member of the genus Vitivirus, family Flexiviridae and was engineered to express the reporter gene beta-glucuronidase (GUS) and the CP gene of citrus tristeza virus in inoculated N. benthamiana plants (Haviv et al., 2006). The GVA vector also was used to silence the endogenous gene in N. benthamiana and Vitis vinifera cv. Prime plantlets (Muruganantham et al., 2009). The vector successfully expressed the reporter genes in N. benthamiana plants and silenced the PDS gene in both N. benthamiana and V. vinifera plantlets (Haviv et al., 2006; Muruganantham et al., 2009). However, this vector was not stable and the inserted sequence was removed from the genome (Haviv et al., 2006). Another GLRaV-2-based vector was utilized to express GFP and knock down the PDS and the magnesium-protoporphyrin IX chelatase (Chl1) genes in V. vinifera plants (Kurth et al., 2012). The vector had the genetic capacity to hold inserts of at least ~2 kb in size and the durability to accommodate the intact inserted cassettes for more than a year (Kurth et al., 2012).
As tombusviruses are well-studied plant viruses in the model plants such as Nicotiana species and have a wide host range, the development of tombusvirus-based vector for grapevine is expected to be a powerful tool for analysis of gene functions and delivery of resistance gene for pathogen control. To our best of knowledge, the GALV-based vector is the only tombusvirus-derived expression and VIGS vector for grapevine.

 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






