


Bacterial blight caused by Xanthomonas oryzae pv. oryzae (Xoo) is a widespread and destructive disease of rice. The yield loss can reach 70% in heavily infected fields, and infection of plants at the seedling stage may result in Kresek syndrome, possibly leading to complete destruction of the crop (Mew et al., 1993; Salzberg et al., 2008). Generally, Xoo enters rice leaves through wounds or hydathodes, and systemically spread through the xylem to other plant parts. The pathogenesis of Xoo is a complex process involving several virulence factors such as exopolysaccharides, extracellular enzymes, adhesins, type 3 secretion system, and motility (Das et al., 2009; Rajeshwari et al., 1997; Ray et al., 2002; Song and Yang, 2010). Although several virulence factors involved in bacterial blight have been discovered, the pathogenesis is not yet perfectly understood.
Bacterial secretion systems are classified into seven groups (Types I-VII) according to their structure, function, and the specific effectors involved (Green and Mecsas, 2016). The type 6 secretion system (T6SS) is a contact-dependent injection system that delivers effector proteins into bacteria or eukaryotic targets (Schwarz et al., 2010). It has been first identified in Vibrio cholerae in 2006 and was then found in most gram-negative bacteria, which use it to translocate effector proteins to target cells (Jani and Cotter, 2010; Leiman et al., 2009; Pukatzki et al., 2006).
T6SS has been reported as a virulence factor in various pathogenic bacteria, such as Pseudomonas aeruginosa, V. cholerae, and Burkholderia pseudomallei (Burtnick et al., 2011; Hood et al., 2010; Joshi et al., 2017). P. aeruginosa exhibited decreased virulence to Arabidopsis thaliana when the expression of hcp, which is a main component of T6SS, was restricted (Lesic et al., 2009). However, not all T6SSs are involved in virulence to eukaryotic hosts, and further research indicated that T6SS plays a key role in the competition between bacteria. The pathogenicity of P. syringae pv. tomato to tomato plants was not affected by defective T6SS, yet, its ability to compete with other Escherichia coli and yeast was highly dependent on T6SS (Haapalainen et al., 2012). Although T6SS has been reported as a virulence factor in other plant pathogens, its role in Xoo is still unknown.
T6SS is structurally homologous to T4 bacteriophage tail proteins (Pell et al., 2009). The essential core component consists of 13 proteins named TssA to TssM (Joshi et al., 2017). The Tss complex forms a basal plate anchored to the inner membrane, and the membrane anchoring complex is comprised of TssJ, TssL, and TssM (Aschtgen et al., 2010). TssB and TssC form the outer sheath, which accelerates the delivery of the inner tube to the target cell (Basler et al., 2012). The inner tube is composed of hexamers of Hcp (hemolysin co-regulated protein). VgrG (valine-glycine repeat protein G), which is located at the tip of the Hcp complex, punctures the outer membrane of the target cell. The key components of T6SS are Hcp and VgrG proteins, which resemble the phage tail and needle complex, respectively (Yang et al., 2018). All T6SS clusters essentially contain Hcp and VgrG proteins, and when these proteins are disrupted, the entire T6SS is dysfunctional.
As T6SS has not yet been examined in Xoo, this study aimed to characterize T6SS in Xoo and to explore its possible involvement in the virulence to host plants and/or in inter-bacterial competition.
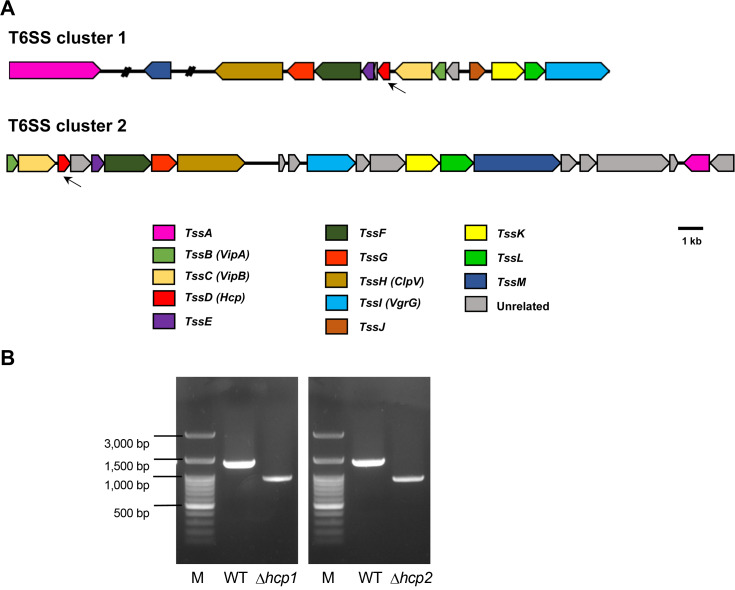
The genome of Xoo PXO99A, the Philippine race 6 strain, which is composed of a single chromosome of 5,240,075 bp, has been previously analyzed (Keshri et al., 2014). The analysis revealed two T6SS gene clusters, T6SS-1 (PXO_02040-PXO_02050, PXO_04696, PXO_04699), containing all the 13 essential proteins of T6SS, and T6SS 2 (PXO_00245-PXO_00266), lacking TssJ, one of the membrane anchoring complexes (Fig. 1A).
In order to study the functions of T6SS-1 and T6SS-2, markerless deletion mutants targeting the hcp gene in each cluster, as a crucial component of T6SS, were constructed. The Δhcp1 mutant is deleted for the 492 bp of the hcp1 gene (PXO_02047) of T6SS-1, Δhcp2 is deleted for the 427 bp of the hcp2 gene (PXO_00264) in T6SS-2, and Δhcp12 is a double mutant deleted for both genes (PXO_02047 and PXO_00264).
The wild-type strain, Xoo PXO99A, and each mutant strain were cultured at 28┬░C with shaking at 200 rpm in nutrient broth and nutrient agar (MBcell, Seoul, Korea), Escherichia coli strains were cultured in Luria-Bertani (LB; Difco, Detroit, MI, USA) agar. For the growth curve, the bacteria were cultured in broth medium at 37┬░C with shaking at 200 rpm, while tryptone yeast extract glucose with 0.6% agar (tryptone, 1 g/l; yeast extract, 0.5 g/l; glucose, 10 g/l; NaCl, 1 g/l; agar 0.6%) was used for the swarming motility assay.
Deletion mutants in Xoo PXO99A were designed using the pK18mobsacB plasmid, which can induce deletion by double-crossover homologous recombination (Sch├żfera et al., 1994). The primers for amplification of fragments upstream and downstream of each deletion mutant strains were designed according to Gibson assembly reaction (Gibson et al., 2010). All primers used in this study are shown in Supplementary Table 1. The upstream and downstream DNA fragment were amplified by PCR, mixed at the same concentration, and re-amplified by secondary PCR using the LF and RR primers of each strain. LF primers contained EcoRI restriction enzyme digestion sites, while RR primers contained HindIII sites. The product of secondary PCR was digested with EcoRI and HindIII (NEB, Ipswich, MA, USA). To generate Xoo with the recombinant plasmids, the wild-type Xoo (recipient strain) and E. coli S17-1 (donor strain), harboring identical recombinant plasmids, were conjugated by the bi-parental mating method (Shen et al., 2002). Properly transformed cells were incubated in NB medium for 2 days, and smeared on peptone sucrose agar (PSA) containing 20 ╬╝g/ml of cephalexin and 10% sucrose (w/v). The obtained deletion mutants were confirmed by PCR assays and sequencing (Fig. 1B). All bacterial strains and plasmids used are listed in Supplementary Table 2. The growth rates and bacterial swarming motility of the obtained mutants were compared to those of the wild-type Xoo PXO99A to evaluate the role of T6SS.
A pathogenicity test was performed to evaluate the involvement of T6SS in Xoo virulence to rice plants. Six-week-old rice (Oryza sativa L. Saeilmi) plants were inoculated by the mutant and the wild-type strains using the scissor clipping method (Mew et al., 1993; Staskawicz et al., 1995). Briefly, each strain was cultured on peptone sucrose broth, and the cells were harvested by centrifugation, washed twice, and re-suspended in sterilized distilled water (SDW). The final OD600 was adjusted to 0.8. At 14 days post-inoculation, the disease severity was evaluated by measuring the lesion lengths. The bacterial population colonizing the rice leaves was assessed by grinding the leaves in SDW, spotting serial dilutions on PSA plates, and expressed as colony forming units (cfu)/leaf.
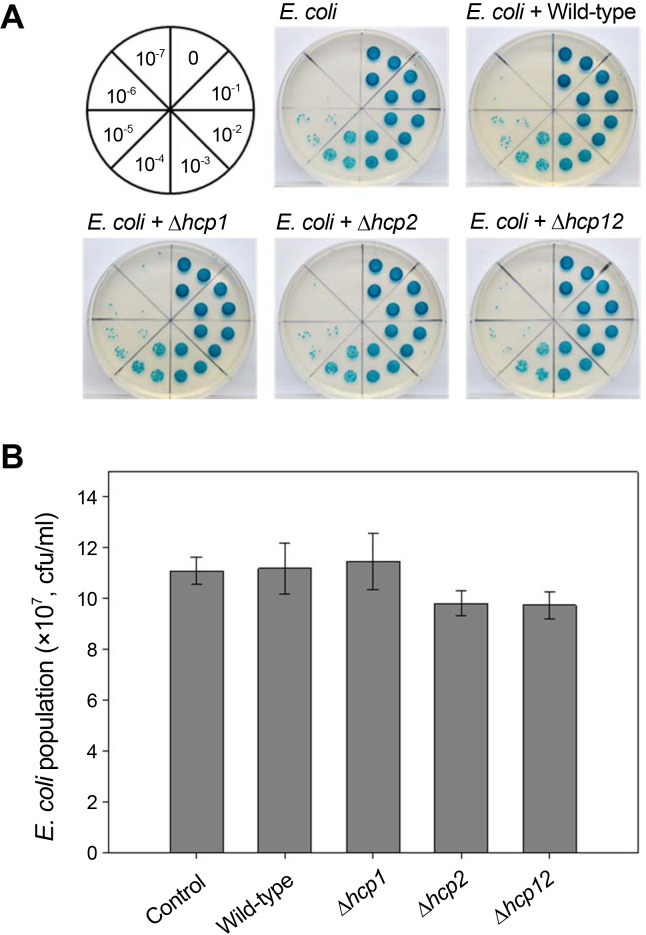
To test whether T6SS was involved in the competition with other bacteria, an inter-bacterial competition assay was conducted using E. coli DH5╬▒ cells as a prey, following a previously described method with some modifications (Ma et al., 2014). The mutants and the wild-type Xoo strains were co-cultured with E. coli DH5╬▒ cells harboring the pCRISPomyces-2 plasmid, allowing for survival in apramycin media and alpha complementation for ╬▓-galactosidase. Bacteria were cultured at a 1:1 ratio in forms of patches on LB plates, and incubated for 12 h. Following re-suspension of the patches, the survival of the E. coli prey cells was assessed after spotting serially diluted patches from 0 to 10-7 on LB plates containing apramycin (100 ╬╝g/ml) and 5-bromo-4-chloro-indolyl-╬▓-D-galactopyranoside (X-gal, 40 ╬╝g/ml). The number of blue colonies on the plate indicated the number of surviving E. coli cells.
The interbacterial competition was also performed using the wild-type and the double mutant Δhcp12 against other bacterial strains as preys (B. glumae BGR1, B. gladioli KACC18962, B. plantarii KACC18965, Ralstonia solanacearum and Burkholderia sp. KJ006). Briefly, Xoo wild-type or Δhcp12 were inoculated into LB gar 0.8% to a final OD600 value of 0.2 that was then poured on LB agar 1.5%. 10 μl from the prey cultures adjusted to OD600 = 0.2 were spotted onto the plates and incubated for 24 h at 30ºC. Bacterial growth was then observed in comparison to the controls.
Statistical analysis was performed using the Statistical Analysis Systems (SAS Institute, Cary, NC, USA). Analysis of variance was performed using the general linear model, and separation of means was determined by Tukey's Honest Significant Difference test, setting P < 0.05 as the statistical significance threshold.
The growth rates and bacterial swarming motility of Δhcp1, Δhcp2, and Δhcp12 mutant strains were not significantly different from those of the wild-type (Fig. 2). These results indicated that neither T6SSs were involved in Xoo growth or motility, which are critical virulence factors in these bacteria (Tian et al., 2015).
When the mutant and wild-type strains were tested for virulence to rice plants, the mutant strains defective in Δhcp2 and Δhcp12 showed a significant reduction in disease severity compared to the wild-type strain (Fig. 3A and B). However, there was no significant difference in disease severity between the Δhcp1 mutant compared to the wildtype strain (Fig. 3A and B). The assessment of bacterial population surviving in rice leaves, expressed as (cfu/leaf), was consistent with the results of disease severity, as the mutant strains, Δhcp2 and Δhcp12, showed significantly reduced survival compared to the control and the Δhcp1 mutant (Fig. 3C). These results implied that Hcp of T6SS-2 was involved in the virulence of Xoo to host rice plants, while no apparent role was associated with T6SS-1. When the complemented mutant Δhcp2C was tested to confirm hcp2 involvement in the virulence to rice, the complemented strain showed recovery of virulence and Xoo population surviving of the leaves comparable to the wild-type (Fig. 3D-F).
According to the inter-bacterial competition assay, there was no difference in the number of surviving E. coli prey cells after co-cultivation with the mutants Δhcp1, Δhcp2, and Δhcp12, as compared to the wild-type (Fig. 4). Additionally, there were no interactions with other pathogenic bacterial spp. (B. glumae BGR1, B. gladioli KACC18962, B. plantarii KACC18965, R. solanacearum and Burkholderia sp. KJ006) as shown in Supplementary Fig. 1. Based on these findings, it could be inferred that T6SS in Xoo is not involved in inter-bacterial competition. However, further research is required to confirm that T6SS has no role in the interaction with other surrounding microbes.
In this study, screening of the Xoo PXO99A genome resulted in the discovery of two T6SS clusters. A deletion mutation of the hcp gene in each cluster was constructed and the obtained strains were tested for virulence to rice plants and for inter-bacterial competition compared to the wild-type strain. The results indicated that the hcp deletion in T6SS-2 partially impaired the bacterial virulence to rice plants, whereas its deletion in T6SS-1 did not result in any significant difference in virulence to rice. This finding was in accordance with previous work on the association of T6SS with the virulence to eukaryotic hosts. Regarding phytopathogenic bacteria, T6SS was reported to be important for the virulence of R. solanacearum to host plants (Zhang et al., 2014).
It has been previously reported that, in some bacterial species, T6SS is involved in the translocation of toxic effectors to other bacteria as a mean of survival competitive advantage (Haapalainen et al., 2012; Russell et al., 2011). However, the inter-bacterial competition assay conducted in this study indicated that disruption of both T6SS clusters did not affect Xoo ability to compete with other bacteria.
The NCBI BLAST analysis demonstrated that Hcp2 in Xoo has a highly conserved amino acid sequence, with respect to the Hcp homolog harbored by X. citri (Xac4145), which is involved in the resistance to the amoeba, Dictyostelium (Bayer-Santos et al., 2018). Therefore, Hcp2 in Xoo might be involved in the interaction with other eukaryotic cells, including amoeba and yeast, in addition to rice plant. The amino acid sequence of Hcp1, however, did not correspond to other Hcp sequences with known functions.
Most T6SSs are strictly regulated at the transcriptional level to ensure the activation of the system only under appropriate conditions (Bernard et al., 2010). T6SS regulation in Xoo has not been studied yet. In V. cholerae, P. aeruginosa and Pectobacterium atrosepticum, the regulation of T6SS is mediated by quorum sensing (Ishikawa et al., 2009; Lesic et al., 2009; Liu et al., 2008). In addition, according to the first relevant study in the genus Xanthomonas, the T6SS of X. citri, was regulated by EcfK, an alternative sigma factor of the ECF family and a transmembrane eukaryotic-like serine threonine kinase (Bayer-Santos et al., 2018).
Since the main role of T6SS is the translocation of various effectors to host cells, it is important to study the secreted effectors. Comparative proteomics analysis in wild-type and T6SS deficient strain could be utilized to identify effectors that are delivered by Xoo to host cells via T6SS (Lien and Lai, 2017; Schwarz et al., 2010).
In conclusion, the present study is the first to functionally characterize T6SS in Xoo and to unveil the role of T6SS-2 in bacterial virulence to rice plants. The presented results are a step toward the complete understanding of the pathogenesis of bacterial blight caused by Xoo, and may facilitate the development of novel disease management options.
Electronic Supplementary Material
Supplementary materials are available at The Plant Pathology Journal website (http://www.ppjonline.org/).


 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement
Supplement Print
Print






