

In Arabidopsis, perception of the bacterial protein, flagellin, occurs at the plasma membrane through the FLAGELLIN-SENSITIVE 2 (FLS2) and BRI1-ASSOCIATED KINASE 1 (BAK1) complex (Gomez-Gomez and Boller, 2000; Chinchilla et al., 2007). FLS2 binds to flg22, a 22-amino acid flagellin-derived peptide that is a pathogen associated molecular pattern (PAMP). Once bound to flg22, the complex initiates several defense responses, including calcium flux, activation of several MAPKs, an oxidative burst, and induction of ethylene and SA biosynthesis. These responses are involved in triggering the intermediate and late defense responses, including activation of a diverse array of defense genes, callose deposition, and inhibition of seedling growth (Boller and Felix, 2009; Tsuda et al., 2008).
MAPK cascades are signaling pathways downstream of sensors or receptors that transduce extracellular stimuli into intracellular responses in eukaryotes. The Arabidopsis MAPKs, MPK3, MPK4 and MPK6, are activated by flg22 treatment within five minutes (Asai et al., 2002; Droillard et al., 2004). Activation of the MAPK cascade positively regulates the expression of several defense-related genes and confers resistance to plant pathogens (Asai et al., 2002). MPK3 and MPK6 are pivotal in systemic acquired resistance (SAR) and SA-mediated priming of plants for disease resistance (Beckers et al., 2009).
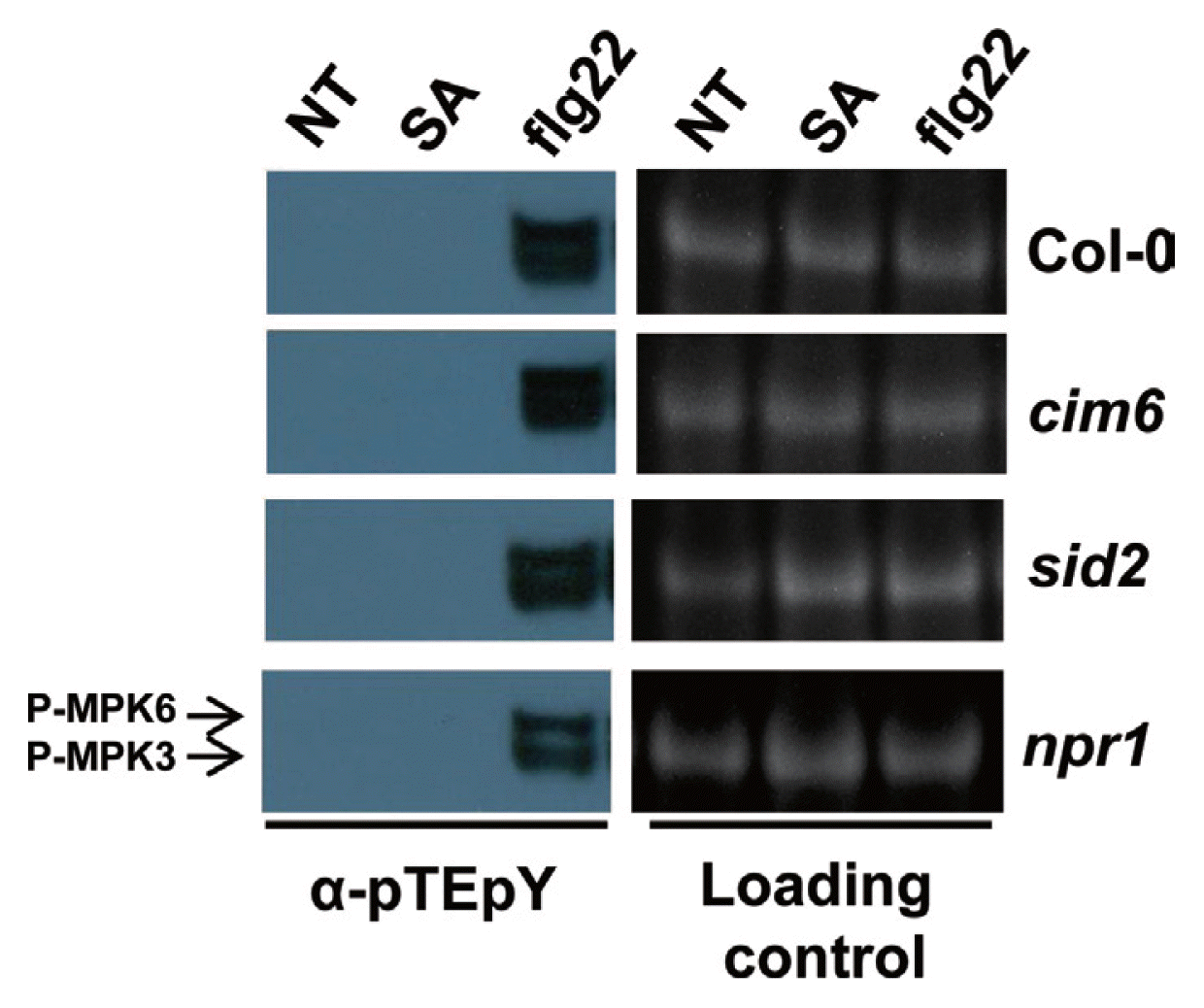
The flg22-triggered oxidative burst and induction of transcription of the gene for WRKY29 and the gene for the flg22-induced receptor-like protein FRK1, which are early responses, have been shown to depend on SA signaling (Yi et al., 2014). We wanted to know if SA signaling also has a role in a very early flg22 response, MAPK activation. We treated wild-type (WT) Arabidopsis seedlings and several SA-related mutants with SA or flg22 and assessed dual phosphorylation of the TEY motif of MAPK. The plants used were WT and the mutants, sid2, which has low SA content (Nawrath and Metraux, 1999); cim6, which has high SA content (Maleck et al., 2002); and npr1, which has a block in SA signaling (Cao et al., 1994). As is shown in Figure 1, when plants were treated with SA, no TEY phosphorylation of MPK3 and MPK6 was detected in any of the seedlings, whereas it was observed after flg22 treatment. Recent report showed that during development of benzothiadiazole (BTH; chemical analogue of SA) induced resistance in Arabidopsis and the priming is associated with accumulation of mRNA and inactive proteins of MPK3 and MPK6 (Beckers et al., 2009). In other words, SA or BTH involved in the accumulation of unphosphorylated MPK3/6 but not in activation of MPK3/6.
It has been suggested that these MAPKs function downstream of reactive oxygen species (ROS) in plant immunity, because they are activated by exogenous H2O2 (Colcombet and Hirt, 2008). Because cim6 and sid2 mutant seedlings are reported to have altered flg22-triggered ROS production (Yi et al., 2014), we expected altered flg22-induced MAPK activation in these mutants. However, all of the mutants exhibited WT-like flg22-induced activation of MAPKs (Fig. 1). Mersmann and colleagues also observed an impaired oxidative burst accompanied by WT-like activation of MPK3 and MPK6 in flg22-stimulated ethylene-insensitive mutants (Mersmann et al., 2010). Based on our results, we suggest that the flg22-triggered oxidative burst and activation of MPK3/MPK6 are independent early signaling events in plant immunity, acting downstream of FLS2.
Flg22 perception by FLS2 leads to the induction of transcription of WRKY29 and FRK1. This induction occurs through the activation of a MAPK signaling cascade consisting of MEKK1, MKK4/MKK5 and MPK3/MPK6 (Asai et al., 2002). Transcript levels of WRKY29 and FRK1 were reduced by approximately 50% in sid2 compared with WT, one hour after flg22 treatment (Yi et al., 2014). However, sid2 showed WT-like flg22-triggered MPK3 and MPK6 activation (Fig. 1). These studies suggest that flg22-induced WRKY29 and FRK1 transcription are regulated by SA signaling as well as activated MPK3 and MPK6 signaling, but two signaling act independently.
Priming with low concentrations of SA or BTH has been reported to increase resistance to subsequent pathogen infection, as evidenced by faster and greater expression of defense genes compared to non-primed plants (Prime et al., 2006). It has also been shown that BTH pre-treatment induced the accumulation of unphosphorylated, inactive forms of MPK3 and MPK6 in Arabidopsis, but when these primed plants were stressed, they accumulated more phosphorylated, active MPK3 and MPK6 than unstressed plants. This enhanced accumulation correlated with greater expression of PAL, which codes for phenylalanine ammonia lyase, and of PR-1, which codes for the pathogenesis-related protein 1 (Beckers et al., 2009). The potentiated responses to pathogen infection, wounding, or infiltration of water into leaves that result from priming with SA or BTH require expression of the NPR1/NIM1 gene in Arabidopsis (Kohler et al., 2002). NPR1 is also involved in SA-induced priming for enhancing flg22 responses, the oxidative burst, the induction of WRKY29 and FRK1 transcription, and callose deposition (Yi et al., 2014). To determine whether NPR1 is also important in priming for enhanced flg22-triggered MAPKs activation, we analyzed the dual TEY phosphorylation in WT and SA-related mutants (Fig. 2). The results show that pretreatment with SA served as an flg22-signaling enhancer in WT, cim6, and sid2 seedlings. The npr1 seedlings, however, did not show SA-dependent enhancement of the flg22-triggered MAPK activation, indicating that NPR1 is instrumental in priming for stronger activation of MPK3 and MPK6. Interestingly, MPK3 activation is stronger than MPK6 in Arabidopsis seedlings treated with SA+flg22. However, both MPK3 and MPK6 seem to be important for full priming in Arabidopsis. This may explain why knockout of any one of the two genes encoding MPK3 or MPK6 doesn’t always attenuate induced resistance (Beckers et al., 2009).
Our results from a previous study (Yi et al., 2014) suggest that SA acts at an early stage of the flg22 response, on the oxidative burst that may be involved in callose deposition, a late flg22 response. Our current results suggest that impaired SA signaling has no effect on the flg22-triggered MAPK activation. It may be that the flg22-triggered oxidative burst and MPK3 and MPK6 activation are two independent, early signaling events in plant immunity, acting downstream of FLS2. Moreover, we provide evidence that NPR1 is involved in SA-dependent priming for enhancing the flg22-induced MPK3/MPK6. Based on previous reports (Beckers et al., 2009; Conrath et al., 2002; Kohler et al., 2002; Yi et al., 2014) as well as on these findings, we propose that NPR1-mediated priming of defense responses is also important in flg22 responses.

 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






