A Review of the Potency of Plant Extracts and Compounds from Key Families as an Alternative to Synthetic Nematicides: History, Efficacy, and Current Developments
Article information

Abstract
The global nematicides market is expected to continue growing. With an increasing demand for synthetic chemical-free organic foods, botanical nematicides are taking the lead as replacements. Consequently, in the recent years, there have been vigorous efforts towards identification of the active secondary metabolites from various plants. These include mostly glucosinolates and their hydrolysis products such as isothiocyanates; flavonoids, alkaloids, limonoids, quassinoids, saponins, and the more recently probed essential oils, among others. And despite their overwhelming potential, variabilities in quality, efficacy, potency and composition continue to persist, and commercialization of new botanical nematicides is still lagging. Herein, we have reviewed the history of botanical nematicides and regional progresses, the potency of the identified phytochemicals from the key important plant families, and deciphered some of the impediments involved in standardization of the active compounds in addition to the concerns over the safety of the purified compounds to non-target microbial communities.
Plant-parasitic nematodes (PPNs) are recognized as important pathogens of agricultural crops worldwide. Annual global economic loss caused by PPNs is estimated to be at US$157 billion (Abad et al., 2008). This figure is most likely an underestimation, as most agricultural farmers are unaware of the existence of nematodes due to their microscopic nature and the more subtle effects of the widely occurring low level infestations (Coyne et al., 2018; Nicol et al., 2011). The symptoms caused by nematode damage are also sometimes difficult to differentiate from those caused by other pathogens and thus, in the absence of nematology experts, such symptoms are conveniently attributed to the common and obvious fungal or bacterial causal agents (Coyne et al., 2018). Additionally, many PPNs do not cause total crop losses. However, the wounds inflicted on the roots during their feeding activity often provide pathways for entry of fungal and bacterial pathogens. PPNs may therefore act in synergy with other pathogens towards reducing crop quality, quantity and causing eventual yield loss (Karssen et al., 2013). Despite the difficulties in nematode damage diagnosis, severe crop losses have evidently been documented depending on nematode species.
In nematode infested cropping systems, control and management is necessary and this involves using integrative approaches (Atolani and Fabiyi, 2020; Lee et al., 2017; Viaene et al., 2013). However, among the available control strategies, nematicides continue to dominate; used either integratively with other management approaches or as the sole control method (Haydock et al., 2013). This is evidenced by the big global market for nematicides worth a billion US dollars per annum, utilizing approximately 350,000 tons of active substance every year (Haydock et al., 2013). With the continuous demand for high quality vegetable crops, the greatest proportion of these chemicals are used in vegetable production, targeting mainly the notorious root-knot nematodes (RKNs). Nematicides have dominated nematode control programs since the late 19th century when fumigants like carbon-disulphide were introduced. They are considered a potent and faster control method against the most important nematode groups in crop production (Karssen et al., 2013; Regmi and Desaeger, 2020). For decades, chemicals like methyl bromide stood out as the standard fumigants for managing nematodes. However, these once-dominant nematicides are no longer available due to their adverse effects in the environment (Regmi and Desaeger, 2020; Sande et al., 2011).
Consequently, more fumigants such as chloropicrin, 1,3-dichloropropene, 1,2-dibromo-3-chloropropane, formaldehyde, and metam sodium among others, were developed in the first and the early second half of the 20th century. And despite the documented side effects, some of them are still in use today (Desaeger et al., 2017; Sande et al., 2011). For example, compounds like 1,3-dichloropropene are under restricted use due to the documented side effects on non-target organisms, and ground-water contamination. Non-fumigants like imicyafos and fosthiazate are being recognized as better alternatives (Kim et al., 2016). But, current markets demand for pesticide/nematicide free agricultural produce.
Botanical pesticides, normally used as plant extracts or soil amendments stand out as one of oldest studied, tested, and available options as detailed by Prakash and Rao (2018) and Yoon et al. (2013). Phytochemicals are generally believed to be environmentally safe, with minimal side effects on non-target organisms. Thus, several plant compounds have been screened for nematicidal or nemastatic properties in addition to the potential use as insecticides and cosmeceuticals, among others (Atolani and Fabiyi, 2020). Currently, substantial progress in the use of phytochemical compounds, in form of plant extract compounds or as whole-plant (biofumigants and amendments) in the management of PPNs is evident, albeit with varying efficacies. Extracts from roots, tree barks, leaves, seeds and fruits have been tested on various important PPNs in crop production (Atolani and Fabiyi, 2020; Mokrini et al., 2018). Herein, we explore a three-decade extensive work and progress on the use of plant extracts and compounds in PPN control. A review on the long history of botanical nematicides and regional progresses in terms of product development, and the potency of the identified phytochemicals from the key important families is given. In addition, we also make sense of some of the impediments arising in standardization of the active compounds, in addition to the concerns over the safety of the purified compounds on non-target microbial communities.
History of Using Plant Materials as Nematicides
The practice of using plant extracts for pest management and control in crop production is not new. It dates as far back as 400 BCE from Rome, clearly portrayed in the meaning of the Lithica poem ‘all the pests that out of earth arise, the earth itself the antidote supplies’ (Dayan et al., 2009). Several scholars dating as far as 234-149 BCE to 23–79 CE detailed various cultural methods of pest management including the use of plant oil extracts. In China, the anthelmintic activity of plants such as Melia azedarach was acknowledged in ancient times dating as early as 25–220 CE. A list of 267 plant species known to possess bioactivity against various crop pests in China by 1950s is detailed (Yang and Tang, 1988). The use of botanical insecticides invigorated in the last millennium in especially China, Egypt, India, United States, Greece and other European countries before the discovery of the fast-acting organochlorine and organophosphate pesticides in the late 1930s and early 1940s (Isman, 1997, 2006; Isman and Seffrin, 2014).
Application in soil were mainly in form of crude extracts or raw plant materials like leaves, seed, oil cakes and whole plant extracts based on the prior knowledge of the cidal effects of various tree species on crop pests (Ellenby, 1945; Sayre et al., 1965). A variety of plants were screened for nematicidal efficacy against the most damaging species as detailed by Prakash and Rao (2018). However, the application of these promising natural products was kept in check by the discovery of the then ground-breaking fast acting organochlorines like DDT, organic halogen compounds (methyl bromide), carbamates and organophosphate nematicides in 1940s and 1950s. Soil fumigants, such as methyl bromide, which contribute to ozone depletion, were the most effective control agents for managing PPNs until their safety profile for the environment and non-target organisms were questioned. It was phased out in 2005 and banned in 2015 under the Montreal Protocol. Similar effective products like 1,3-dichloropropene are now banned in the EU and are under restricted use in most developing countries (Haydock et al., 2013). This prompted the development and use of organophosphates such as fenamiphos and fosthiazate, and carbamates like carbofuran, and aldicarb in second half of the 20th century. However, most of these nematicides are evidently groundwater contaminants and are persistent in the soil. This has resulted in either restrictions in use or eventual withdrawal from the chemical market (Fishel, 2006; Ruiz-Suárez et al., 2015), hence amplifying the need to screen less toxic and environmentally friendly substitutes.
Consequently, many researchers have been attempting to screen and develop phytochemical-based nematicides based on the chemical interactions between nematodes and plants. A number of the isolated plant compounds involved in plant-nematode interactions are categorized among nematode attractants, repellents, hatching inhibitors or stimulants, and toxicants (Chitwood, 2002; Ntalli and Caboni, 2012). These interactions, which are part of the ever-growing discipline of allelopathy, have received greater attention during the past two decades. The screened compounds are often a mixture of biologically active substances, an attribute that reduces the possibilities of resistance development by the target nematodes (Dimetry, 2014; Saxena, 1983). The use of botanical nematicides has therefore been widely recommended as a suitable alternative of plant protection, with minimal side effects though there are some documented reports of undesirable effects on non-target microbial communities (Caboni et al., 2012a; Isman, 2006; Miresmailli and Isman, 2014). On the other hand, it is also important to note that many of these plant materials are often slow-acting; and the economic cost of screening and the eventual commercial production presents enormous doubt on the sustainability of scaling up production of some of the tested compounds. Thus, many of the investigated plants, especially in the developing world continue to be used as organic manure or residues incorporated in soil (Chitwood, 2002; Ntalli and Caboni, 2012). Nevertheless, there is a large volume of phytochemical screening investigations on botanical nematicides, and commercialization of these new botanical compounds have seen marked increase in recent years (Grieneisen and Isman, 2018). Registration of botanical compounds has seen more success in the US, due to regulatory exemptions in the commercialization process of some essential oil (EO)-based pesticides, unlike regions like Europe, where regulatory procedural exemptions are not granted, and commercialization still lag behind (Caboni and Ntalli, 2014). Identification of active ingredients is currently relatively easy due to the ever-growing advances in analytical procedures and this is evidenced by a substantial increase in the proportion of published research over the past 30-years.
Recent and Current Regional Developments
In the last three decades, the interest and use of botanical nematicides has been on a steady increase worldwide. In the current review, a large volume of phytochemical screening investigations was uncovered. Regionally, Asia is taking the lead in research on botanical nematicides (35% of the published screening studies), owing to the richness of medicinal plants in the region. Africa and Europe represent 23 and 21%, respectively, North and South America, 17%, while Oceania represents 1.7%. However, a significant proportion of active compounds remain unidentified in a large volume of the screened plant materials in Asia (23%) and Africa (33%), compared to North America and European countries. In North America (US) for example, almost all the screened plant materials have their active compounds identified, and registration of new EO-based biopesticides is widely favored (Caboni and Ntalli, 2014) (Table 1).
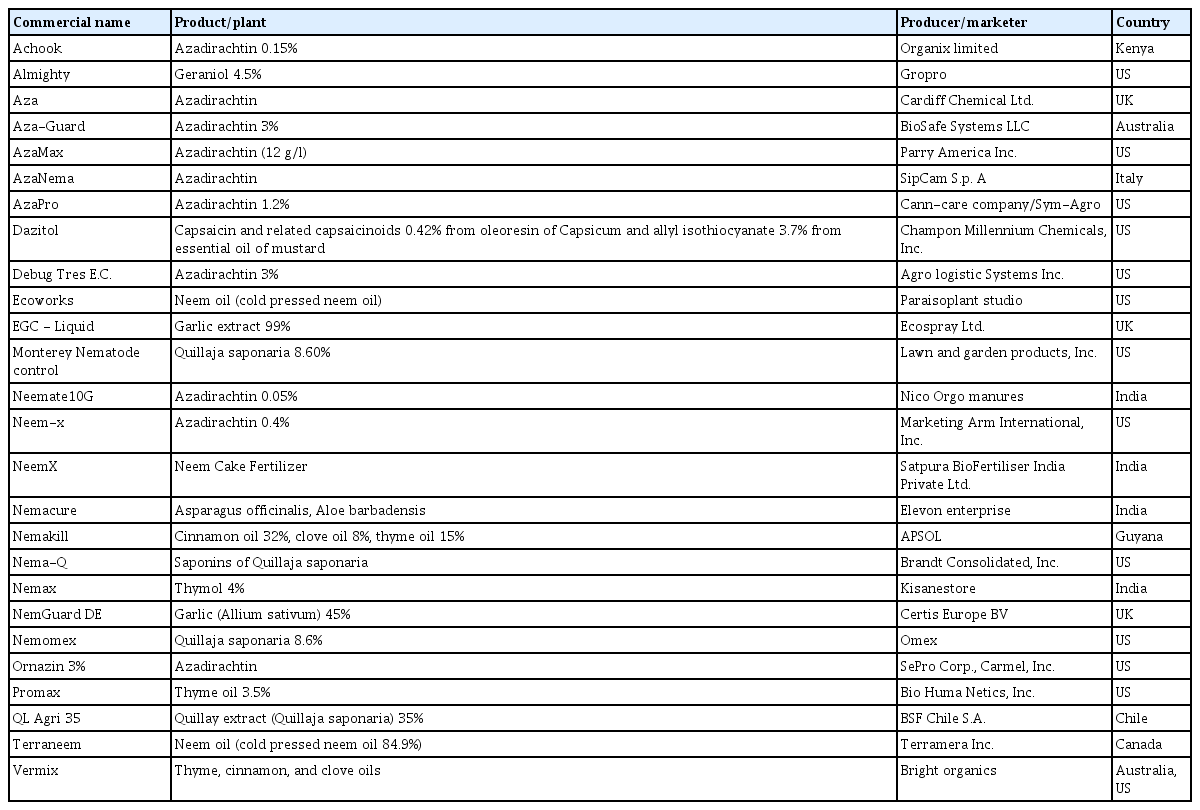
Examples of available commercial botanical nematicides, with their respective active constituents
Over 348 plant species belonging to over 81 families have been screened for nematicidal bioactivity. Of the 348, the most dominant species (ca 42% of the published studies) are; Azadirachta (A. indica), Melia (M. azedarach), Eucalyptus spp., Ricinus (R. communis), Satureja spp., Solanum spp., Allium spp., Thymus spp., Citrus spp., and Artemisia spp., and of these, Azadirachta (A. indica) accounts for ca 16% (Fig. 1A). Over 50% of the tested plant species belong to the families; Asteraceae, Fabaceae, Lamiaceae, Brassicaceae, Myrtaceae, Euphorbiaceae, and Apocynaceae; and of these, Asteraceae and Fabaceae represents 30 and 20% of the group (Fig. 1B). Research on these botanical materials also highlights the importance of the most destructive PPN species. Meloidogyne spp. stand out as the main target (80% of the targeted nematode species).
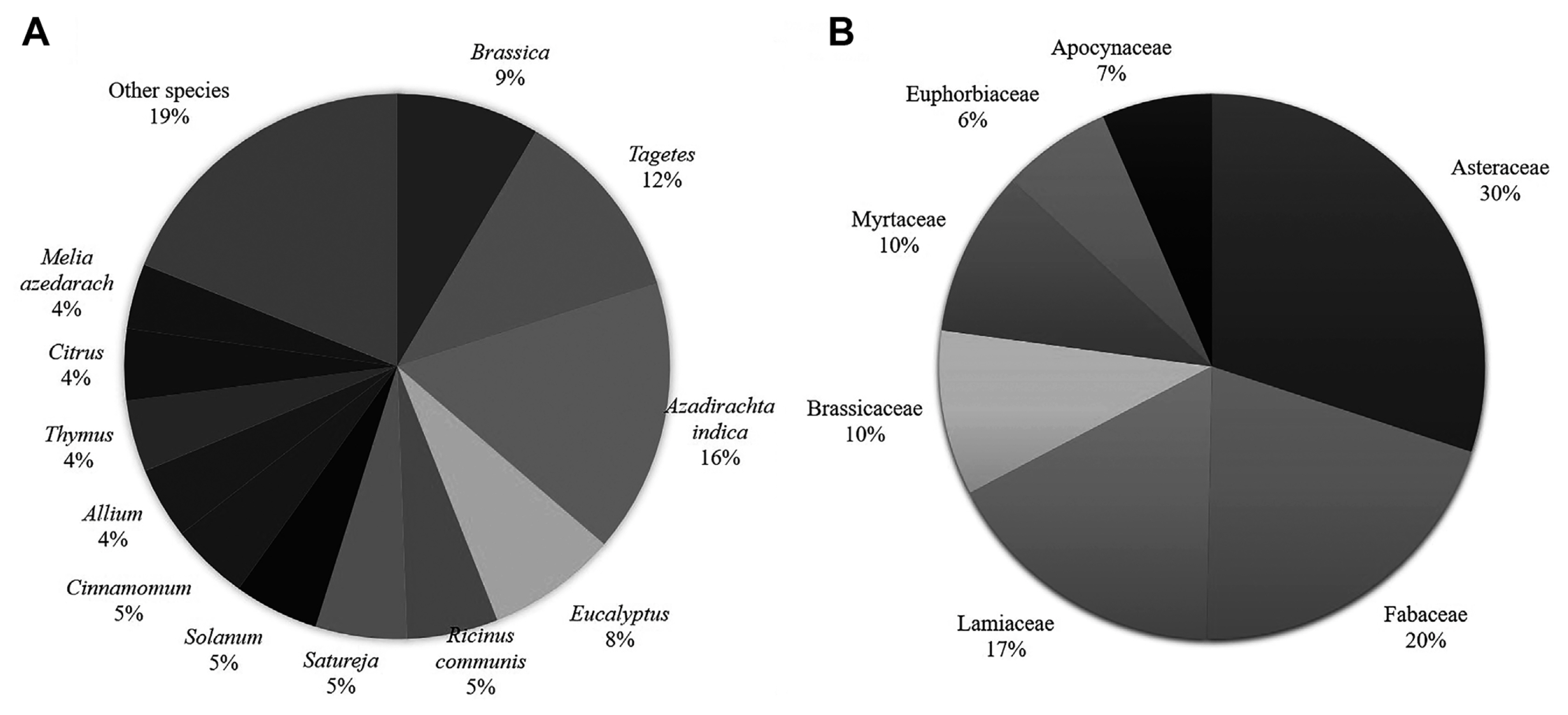
A representation of the most studied plant species and families: (A) 42% of all the most screened species, (B) 50% proportion of the most screened families.
Composition, Activity, and Potency of the Active Constituents
The basis of research on interactive effects of phytochemicals to the target organisms is rooted in allelochemistry; the study of effects of organic compounds, mostly ‘secondary metabolites’ synthesized by plants, and released upon other organisms separated from their sources in the environment (Gross, 2009; Harlin and Rice, 1987). These allelochemicals play ecological roles in counteracting external biotic and abiotic stressors (Ntalli and Caboni, 2012). In this case, there are chemical-mediated interactions between phytochemicals and the target nematodes. In the last 2–3 decades, there have been vigorous efforts towards the identification of the active secondary metabolites from various plants of nematicidal importance. These are mostly glucosinolates and their hydrolysis products such as isothiocyanates; phenolics like flavonoids and their glycosides; aldehydes and ketones; alkaloids; limonoids, quassinoids, and saponins; polyacetylenes and polythienyls; piperamides; organic acids; and the more recently probed EOs (Andrés et al., 2012; Avato et al., 2013; Ntalli and Caboni, 2012; Ntalli et al., 2010a; Park et al., 2007).
Research on nematode control has therefore taken a paradigm shift from the traditional cultural, plant-material fumigation methods, and synthetic chemical control measures to innovations involving specific plant compound annotations, with the prospect of industrialized production and commercialization of safer botanical nematicide formulations. Significant attention is currently given to research on the efficacy of EOs against the most notorious nematode species. EOs are a mixture of natural volatile complex compounds found in various groups of aromatic plants. They are typified by their characteristic odors, and are distinctly a complex mixture of primarily terpenes (especially monoterpenes, sesquiterpenes, diterpenes and triterpenes), terpenoids, and an array of other aliphatic and aromatic constituents (Aharoni et al., 2005). EOs are therefore part of the complex plethora of plant secondary metabolites, and are an important defense strategy against pathogens and insect pests. Being heterogeneous mixtures, the bioactivity of EOs is sometimes synergistically tied to these constituents in a very intricate manner; the final effect being either complementary or antagonistic in nature (Andrés et al., 2012; Ntalli and Caboni, 2012). An assortment of EOs of nematicidal importance have been extracted and annotated from various plant species belonging to different families as listed by Andrés et al. (2012). Their chemical composition varies significantly between plant species, varieties, and even within the same varieties separated geographically. Composition also varies with plant age at the time of extraction, percentage humidity of the harvested plant material, and the extraction method used (Andrés et al., 2012; Lahlou and Berrada, 2003). EOs from families like Lamiaceae, Lauraceae, Myrtaceae, Geraniaceae, Rutaceae, Poaceae, Asteraceae, Fabaceae, Brassicaceae and many others have exhibited effective control potential against root-knot, cyst, lesion and pine wood nematodes, with several of them already scaled up for commercial production (Table 1).
However, whereas reports show that isolated EOs and other principal active compounds in plant extracts exhibit very effective inhibitory and nematicidal activity, it has also been demonstrated that some of these compounds do not only adversely affect the soil microbial community but also exhibit variations in efficacy (Caboni and Ntalli, 2014; Caboni et al., 2012a; Miresmailli and Isman, 2014; Ntalli et al., 2018, 2020). There are therefore concerns with the standardization of the active ingredients in botanical nematicides. For example, Watson and Desaeger (2018) demonstrated that the application of two botanical nematicides of Dazitol (capsaicin and related capsaicinoids 0.42% from oleoresin of Capsicum and allyl isothiocyanate [ITC] 3.7% from EO of mustard) and Nemakill (cinnamon oil 32%, clove oil 8%, thyme oil 15%) did not enhance crop establishment or suppress Belonolaimus longicaudatus populations. Javed et al. (2008) also demonstrated that the exposure of M. javanica to Aza (Azadirachtin) caused neither mortality of infective juveniles (J2s) nor hatching inhibition. However, neem crude extracts were lethal to J2s and inhibited hatching. Similarly, AzaNema (Azadirachtin 1%) and Azadirachtin technical grade were shown to not adequately control M. incognita at the recommended or even higher doses (Ntalli et al., 2009). In the experiments performed by Caboni et al. (2013), Mentha aqueous extracts were more active against M. incognita than isolated EOs. Ntalli et al. (2018) also demonstrated that similar suppression levels could be obtained when fruit extracts or fruit powder of M. azedarach were used in comparison to its major active constituent (furfural) when applied singly. In addition, not only did furfural display phytotoxic tendencies, but also had adverse effects on the soil microbial community; tendencies which are avoidable when semi-refined extracts or powders are used. Crude plant powders and their unfiltered extracts are chemically complex, and have been shown to consistently offer active nematicidal activity with minimal toxic effects to non-target microbial community, in addition to stimulation of nematode antagonists (Ntalli et al., 2017, 2020). The extracts and powders when amended in soil do not only release the active compounds, but are also broken down to produce nitrogenous compounds which are toxic to the target nematodes, in addition to stimulating an increase in beneficial microbial populations in the soil.
Notably, Oro et al. (2020) tested some EOs from several plant families against the free-living nematode Panagrolaimus sp. However, the trend of nematicidal bioactivity was close to that exhibited to PPNs; with EOs of Origanum vulgare, Mentha spicata, M. piperita, Salvia sclarea, and Thymus vulgaris exhibiting significantly low LC50/24h of 0.508, 0.505, 0.405, 0.430 and 0.391 μl/ml, respectively. Ntalli et al. (2020) also noted that whereas the EO of Thymus citriodorus was effective in arresting the biological cycle of RKNs, it showed detrimental tendencies against bacterial and fungal feeding nematodes. However, thyme powder and unfiltered extracts were found to instead augment the bacterial biomass and the free-living nematode populations.
One of the main bases of renouncing the continuous use of synthetic nematicides has always been toxicity against the non-target organisms in the environment. Plant-based nematicides are touted to be a safer alternative. However, in these comparative studies, there is evidence to the contrary, more so when the active ingredients are isolated and singly applied in soil. Currently, little attention is given to the ecotoxicological studies of biopesticides, so long as biological activity against the target organism is assured. Registration and production regulations are even relaxed in some regions, owing to the natural origin of the active compounds. Botanical nematicides are distinct from synthetic pesticides by their complex chemical constitution of often closely related secondary metabolites (Miresmailli and Isman, 2014). These metabolites often act synergistically to augment their effects and thus, play an important role in not only the activity against the target organisms, but also the eventual toxicity levels of the mixtures exhibited against non-target organisms, and the stability of the active constituents, unlike synthetic compounds which are generally formulated as single active compounds laced in inert ingredients (Miresmailli et al., 2006). Currently, active compound extraction and eventual application is relatively easy. However, as more active compounds are singly isolated and scaled up for commercial production, the necessity to compulsorily integrate inclusive biota safety study profiles of these purified active compounds on non-target soil communities should be considered.
The potency of some important families
Current trends in the use of botanical nematicides indicate that most of the investigated allelopathic compounds that exhibit nematicidal, nemastatic, hatching inhibition, and root-galling reduction properties, among others, are largely from member species of the families; Asteraceae, Fabaceae, Lamiaceae, Brassicaceae, Myrtaceae, Meliaceae, Solanaceae, Rutaceae, Amaryllidaceae, Quillajaceae, Euphorbiaceae and several others. Below is a summary of the potency of each group.
The potency of Brassicaceae
Several species of Brassicaceae form the ideal group of plants that have been mainly utilized for decades as organic biofumigants (Matthiessen and Kirkegaard, 2006; Kruger et al., 2013). Brassicaceae have been applied in soil in various forms, including as cover-crops, rotation crops, trap crops, green manure, seed meal and bran, and aqueous leaf extracts (Fourie et al., 2016). The biofumigant properties of this group are owed to the presence of glucosinolates within their tissues; natural components which are easily hydrolyzed by endogenous myrosinase during decomposition to yield compounds like ITCs, ionic ITCs, nitriles and organic cyanides, which are bioactive against PPNs (Cole, 1976). These compounds especially ITCs and ionic ITCs are volatile, and toxic to a wide range of soil borne pathogens; and therefore, mimic the biofumigation properties of methyl ITC, the active ingredient in metam sodium (Matthiessen and Kirkegaard, 2006). However, the compounds are fully biodegradable, less persistent in soil and less toxic to non-target microbial community; and therefore, using Brassicaceae in biofumigation is considered an environmentally friendly substitution to the persistent, yet toxic fumigant compounds such as metam sodium, dazomet and the recently phased out methyl bromide (Vig et al., 2009).
A large number of pure ITCs have been tested against PPNs and are now under production (Dahlin and Hallmann, 2020; Wood et al., 2017; Wu et al., 2011; Zasada and Ferris, 2003). The efficacy of these compounds varies depending on the chemical type, concentration, application period, soil type, and the target nematode species, among other factors. For example, significantly low LC90 of 0.01–0.05 μmol/ml was recorded when pure allyl, benzyl, 2-phenylethyl, and 4-methylsulfinyl(butyl) ITCs were tested against Tylenchulus semipenetrans in sand. Benzyl and 4-methylsulfinyl(butyl) ITCs were also highly toxic to M. javanica, with LC90 values of 0.06 and 0.03 μmol/ml, respectively (Zasada and Ferris, 2003). These toxicity levels were higher when compared to metam sodium (0.08 and 0.13 μmol/ml for T. semipenetrans and M. javanica, respectively). A similar trend was reported by Wu et al. (2011), despite the higher concentrations required to cause mortality of M. javanica in the in vitro test results. Allyl, acryloyl, ethyl, benzyl, 1-phenylethyl, 2-phenylethyl ITCs and benzyl thiocyanate showed irreversible nematicidal activity against J2s of M. javanica after a 72-h exposure to concentrations of 5 μg/ml. And at as low as 0.5 ml/kg soil of allyl and acryloyl ITCs concentrations, the efficacy was equivalent to that of metam sodium at the recommended dose. Allyl ITC was also shown to be very effective at suppressing Globodera pallida through suppressed hatching and egg viability (Wood et al., 2017). Biofumigation using purified natural ITCs is therefore a promising replacement to the more toxic synthetic fumigants.
Nevertheless, members of this family have mainly been utilized as soil amendments. They are known to effectively manage the major PPN groups of economic importance when appropriate plant species such as Brassica oleracea, B. juncea, B. napus, B. rapa, Eruca sativa, Sinapis alba, S. arvensis, and Raphanus sativus are amended in the soil. Sinapis alba seed meals exhibited 100% suppression levels of M. incognita and Pratylenchus penetrans, when applied at a rate of 2.5% and 10% w/w in dry soil. Brassica juncea seed meal was even more effective at a much lower application rate of 0.5%, exhibiting a 100% suppression effect to both species (Mazzola et al., 2009; Zasada et al., 2009). In studies done by Meyer et al. (2011), a 0.2% B. juncea, and 0.05% S. alba + 0.15% B. juncea seed meals, did not only consistently suppress M. incognita population but also, were non-phytotoxic to pepper and lettuce compared to other application rates. Similarly, Lazzeri et al. (2009) reported similar results when B. carinata seed meal was applied to suppress M. incognita on zucchini in a greenhouse.
However, despite the promising potential, inconsistent results have been evident in field applications. For example, in field trials conducted by Engelbrecht (2012), the four tested Brassicaceae cultivars (B. juncea cv. Nemat; E. sativa cv. Calienté; R. sativus cv. Terranova and Doublet) were instead found to either maintain or even increase population levels of M. incognita; Vervoort et al. (2014) also literally demonstrated a no effect on the targeted nematode populations (Tylenchorhynchus spp. and Trichodorus spp.) after a field biofumigation with four B. juncea cultivars (Terrafit, Terratop, Terraplus and ISCI-99); Lord et al. (2011) demonstrated that out of the 22 brassica accessions tested against G. pallida, only 12 significantly inhibited the motility of infective J2s; and when incorporated into the soil containing encysted eggs of G. pallida, only 3 B. juncea lines (Nemfix, Fumus, and ISCI99) known to contain high concentrations of 2-propenyl glucosinolate showed a high efficacy, causing over 95% mortality of encysted eggs. However, the maximum toxic effects were only evident when the soil was covered with polyethylene after the application of green manures. The types and amounts of glucosinolates, and their breakdown products differ depending on the plant species. Therefore, efficacy and phototoxicity possibilities vary depending on the plant species, cultivars, and application rates (Hansson et al., 2008; Mazzola et al., 2007; Meyer et al., 2011). The formulations and particle size for example, has a pronounced influence on the efficacy of seed meals. Zasada et al. (2009) demonstrated that the efficacy of S. alba against P. penetrans improved by 47–56% when the seed meal was ground to smaller particle size, compared to when applied as a pellet.
The potency of Asteraceae
Many allelochemicals including EOs with nematicidal potency have been isolated from several species of Asteraceae (Chitwood, 2002; Oka et al., 2000). The marigolds (Tagetes spp.) represent one of the oldest, and most widely studied plant groups in nematode management. Species like T. erecta (African marigold), T. patula (French marigold), and T. minuta (South American marigold) are effective in the management of nematodes (Bridge, 1996). Marigolds are mainly applied as non-hosts or poor hosts, trap crops, and enhancers of nematode antagonistic flora and (or) fauna. These approaches are either applied separately or in combination, resulting in a synergistic effect on control of PPNs (Akhtar and Malik, 2000; Pudasaini et al., 2006). Several endo-root bacterial isolates like Microbacterium esteraromaticum and Kocuria varians isolated from T. erecta and T. patula roots have also been shown to possess activity against root-lesion nematodes (Sturz and Kimpinski, 2004).
Key nematode groups such as Meloidogyne and Pratylenchus are reported to be consistently suppressed by marigolds. For example, in plots amended with seedlings of African marigold, significant reduction in population densities of M. incognita, Helicotylenchus spp., and Dolichodorus spp. were achieved, with corresponding increase in total N, available P and exchangeable K (Ogundele et al., 2016). Tagetes is known to increase nematode-trapping fungi or parasitic fungi, and enhance both nematode egg and motile form parasitism. In marigold varieties which are resistant to root-knot and lesion nematodes, nematode development is arrested (Hooks et al., 2010; Ploeg and Maris, 1999). The root diffusates of T. minuta are known to inhibit nematode hatching while T. patula instead enhances hatching of for example, P. penetrans; thus, acting as a “dead end” trap crop since it’s a non-host (Evenhuis et al., 2004; Hooks et al., 2010; Pudasaini et al., 2006). However, all these outcomes rely on the abundance of released allelopathic compounds that are toxic, repel or inhibit normal nematode development.
Tagetes spp. are known to contain allelopathic sulphur containing compounds like α-terthienyl and bithienyl which have been reported to possess nematicidal and insecticidal properties (Hooks et al., 2010; Morallo-Rejesus and Decena, 1982; Uhlenbroek and Bijloo, 1958, 1959). Compounds such as thiophenes: 5-(3-buten-1-ynyl)-2,2′-bithienyl (BBT), 5-(4-hydroxy-1-butynyl)-2,2′-bithienyl (BBTOH) and 5-(4-acetoxy-1-butynyl)-2,2′-bithienyl (BBTOAc), including the recently studied EOs have been isolated from various cultivars of T. patula, T. erecta, and T. minuta (Hooks et al., 2010; Szarka et al., 2006). The genus is therefore recognized as a ready source of nematode bioactive chemical constituents, including acetylenes, alkaloids, flavonoids, carotenoids, thiophenes, terpenoids, citric and malic acids (Faizi and Naz, 2002; Saleem et al., 2004; Vasudevan et al., 1997). In the bioassay studies, extracts of flowers of T. patula (rich in α-terthienyl, and flavonoids plus other phenolic compounds like patuletin, quercetin and patulitrin) were shown to exhibit 50–100% mortality of Heterodera zeae after 24-h treatment at 5% concentration (Faizi et al., 2011). EOs z-β-ocimene and dihydrotagetone isolated from T. minuta were shown to be toxic to eggs and J2s of M. incognita; causing J2-hatch inhibition of up to 79% at 4% application rate, and J2 mortality of 100% after 96h (Adekunle et al., 2007). In addition, several other constituents of EOs (limonene, (Z)-β-ocimene, dihydrotagetone, (E)-tagetone, (Z)-tagetone, (Z)-ocimenone, and (E)-ocimenone), have been characterised from T. minuta. These were nematicidal against J2s of M. javanica, with LC90/72h of 0.24–0.45 mg/ml, depending on the cultivar of T. minuta (Massuh et al., 2017).
Other species of Asteraceae such as Stevia rebaudiana, Artemisia annua, Verbesina encelioides, Tithonia diversifolia, Bidens pilosa, Chrysanthemum cinerariaefolium, Chrysanthemum coronarium, C. segetum, Aster sedifolius, A. caucasicus, Achillea wilhelmsii, Ruta chalepensis, Gochnatia barrosii, Heterotheca inuloides, Eclipta prostrata, Vernonia amygdalina, among others are also a source of compounds known to have effective potency against PPNs (Caboni and Ntalli, 2004; Claudius-Cole et al., 2010; de Santos Júnior et al., 2010; Di Vito et al., 2010; Kepenekçi et al., 2016; Kim et al., 2016; Oka, 2012; Pérez et al., 2003; Taba et al., 2008). Stevia rebaudiana is mainly known for its sweet-tasting glycosides of steviol and is used as natural non-caloric sweetener (Karaköse et al., 2015; Kinghorn and Soejarto, 1985). However, it is also rich in compounds like phenols, terpenes, flavonoids, coumarin based analogues, amino and fatty acids, which have been proved to be nematicidal against RKNs. Specific bioactive compounds known to have chemotactic repulsive properties such as 6-isopropenyl-4,8a-dimethyl-1,2,3,5,6,7,8,8a-octahydro-napthalen-2-ol, flavonol (morin), and the kaempferol derivative, robinin, have recently been identified in S. rebaudiana extracts (Chin et al., 2018; Karaköse et al., 2015). The acetonic extract and some cadinanes of species like Heterotheca inuloides have also been shown to inhibit hatching and immobilize J2s of Nacobbus aberrans (Rodríguez-Chávez et al., 2019).
Artemisia annua aqueous extract, rich in caffeic and chlorogenic acid, artemisinin and artesunate, possesses nematicidal bioactivity against G. rostochiensis and Xiphinema index. D’Addabbo et al. (2013) showed that the aqueous extract was effective against G. rostochiensis, causing a 70% J2 mortality after a 24-h exposure to concentrations of 125 μg/ml. On the other hand, caffeic acid was highly toxic to X. index, causing a 100% mortality at all tested concentrations after 2 h of exposure. Verbesina encelioides extracts and fresh parts’ powder have been tested against M. incognita, M. hapla, M. javanica, Ditylenchus dipsaci, P. mediterraneus, and Tylenchulus semipenetrans. The aqueous extract of the flower is said to possess a more significant nematicidal effect against M. javanica, as J2 exposed to the flower extracts at concentrations >3.13% did not recover after 3 days of exposure. Additionally, J2 hatching was inhibited by leaf and flower extracts when treated at concentrations of 3.13 and 1.6%, respectively (Oka, 2012). Tithonia diversifolia extracts contain significant amounts of alkaloids and saponins, and were shown to inhibit hatching of M. incognita J2s. Its residual treatments at the rate of 30 tons/ha on yam significantly suppressed M. incognita population and root-galling (Odeyemi and Adewale, 2011). Furthermore, Di Vito et al. (2010) demonstrated that by using leaf meal of A. caucasicus, reproduction of M. incognita was suppressed by 82.3 and 92.7% at an application rate of 0.5 and 1 g/100 cm3 soil, respectively. Aqueous extracts of B. pilosa are also said to have significant immobilization, repellence, and lethality effects on M. incognita (Taba et al., 2008).
EOs from Chrysanthemum spp. and Eupatorium spp. have strong nematicidal activity against RKNs. For example, Pérez et al. (2003) demonstrated that EOs from C. coronarium significantly reduced hatching of M. artiellia in the in vitro studies; and a significant reduction in reproduction (83.0–95.9%) was observed when infested soils were amended with flowers, leaves, roots or seeds of C. coronarium, C. segetum, Calendula maritima, C. officinalis, and C. suffruticosa. Additionally, Sosa et al. (2012) demonstrated that the EOs from E. viscidum, rich of compounds like spathulenol and 6-methyl-heptenone have nematicidal effects on M. javanica with an ED50/72h value of 0.1 mg/ml. Vernonia colorata is known to contain significant amounts of flavonoids, condensed tannins and phenolic contents. Its extracts were shown to effectively prevent M. incognita J2 root-penetration and the eventual root-gall formation when egg masses were pre-exposed to 0.8 mg/ml plant extracts for 72 h prior to pot inoculations (Sithole et al., 2021).
Echinacea is another group of plants within the family that are known to contain important bioactive compounds. These include pyrrolizidine alkaloids, glycoproteins, alkamides, polyacetylenes and polyenes, caffeoylquinic and caffeoyltartaric esters, caftaric, cichoric and chlorogenic acid, and echinacoside (Laquale et al., 2020; Pellati et al., 2004, 2006). Extracts from Echinacea spp. (E. angustifolia, E. purpurea, and E. pallida) caused a 50% M. incognita J2 mortality after a 48-h exposure to concentrations of 500 μg/ml. Suppression of nematode multiplication was also evident when Echinacea plant biomasses were emended in M. incognita infested soils (Laquale et al., 2020).
The potency of Meliaceae
The family Meliaceae has received much recognition, owing to the abundance of limonoid triterpene compounds in aerial and below ground plant parts of its species (mainly A. indica and M. azedarach). Neem (A. indica) has by far been widely studied for its predominantly tested tetranortriterpenoid limonoid, azadirachtin. Neem is normally applied as oil cakes, crude plant extracts, root-dipping and seed-coating agents, including coating on fertilizers like urea or whole plant material amendments in soil (Akhtar, 2000; McSorley, 2011). Its refined product azadirachtin is one of most commercialized botanical nematicides (Table 1). Azadirachtin affects the target organism by inducing feeding deterrent effects on chemoreceptors; disruption of endocrine system; influence on behavior and reproduction through direct effects on tissues (Abbasi et al., 2005; Mordue (Luntz) and Blackwell, 1993). In addition to azadirachtin, 100–300 different limonoids and other compounds have been identified in neem, exhibiting nematode and insect antagonistic attributes like deterrence of feeding, repellence, and hatching and growth inhibition (Aarthy et al., 2018; Khalil, 2013; Schmutterer, 1990). Limonoids such as salannin, salannolacetate, 3-deacetylsalannin, nimbin, 6-deacetylnimbin, 6-deacetylnimbinene, nimbinene, nimbanal, azadiradione, azadirone, nimocinol, epoxyazadiradione gedunin, and vilasinin, among others, have been identified and investigated in a wide range of biological and medicinal studies. In addition, other compounds like amino acids, polysaccharides, flavonoids, coumarins, dihydrochalcones, tannins and aliphatic and sulphureous compounds are all part of the constituents of neem chemistry (Aarthy et al., 2018; Biswas et al., 2002; Devakumar and Dev, 1996; Kraus, 1995).
Successful nematode management approaches involving use of neem products have been widely documented. For example, urea coated with nimin, which is a neem-based product was shown to not only significantly reduce the development of RKNs but also improve plant growth (Wani and Bhat, 2012); neem leaves applied at a rate of 30 g/kg soil were found to effectively suppress root-galling and improve plant growth parameters of tomato (Yadav et al., 2018); undiluted standard neem extracts showed 100% inhibition of egg production, reduced root-galling by 85%, and significantly improved the activities of resistance-related enzymes phenylalanine ammonia lyase, polyphenol oxidase and peroxidase (Nile et al., 2018), and neem derived product (Ornazin) strongly reduced nematode reproduction and root-galling in tomato when applied as a drench (López-Pérez et al., 2011). Also, in greenhouse trials, 1% neem cake caused a 67–90 % population reduction of P. penetrans and M. hapla in tomato roots (Abbasi et al., 2005).
Melia azedarach is also known to exhibit strong nematicidal, insecticidal, antifungal and antihelmintic properties (Akhtar et al., 2008; Carpinella et al., 2003; Maciel et al., 2006). Extracts from fruits of M. azedarach are reported to exhibit strong biofumigant properties against M. incognita when incorporated as a powder in soil, owing to the presence of carboxylic acids and aldehydes, and particularly furfural, which is a strong nematicide (Ntalli et al., 2010b). Furfural has been reported as the most active component, with a EC50/1d of 8.5 mg/l (Caboni and Ntalli, 2014). Limonoids like meliacin, meliacarpin, melianol, melianone, meliartenin and 21-β-acetoxymelianol have also been isolated from fruits of M. azedarach (Carpinella et al., 2002; Lavie and Jain, 1967; Lee et al., 1991; Ntalli et al., 2010b). In field studies, Cavoski et al. (2012) reported that crushed fruits of M. azedarach, when applied in the soil against M. incognita at application rates of 30 and 60 g/kg, exhibited nematicidal efficacy similar to that of fenamiphos, in addition to elicitation of cucumber plant defense.
The potency of Lamiaceae
A large number of EOs with nematicidal properties have been extracted and annotated from several species of Lamiaceae (Andrés et al., 2012; Ntalli et al., 2010a, 2020). EOs from this group usually comprise mixtures of terpenes and terpenoids blended with other aliphatic and aromatic ingredients (Ntalli et al., 2010a). Isomers of 1,2-epoxymenthyl acetate, L-carvone, piperitone, pulegone, (−)-carvone, trans-anethole, eugenol, borneol, geraniol, limonene, γ-terpinene, o-cymene, carvacrol, 1,8-cineole, α-pinene, camphor, terpinen-4-ol, eucalyptol, ã-terpinene, methyl eugenol, estragole, and thymol are some of the most important EOs isolated from Lamiaceae, and are considered to have strong nematicidal potency (Andrés et al., 2012; Avato et al., 2017; Barbosa et al., 2010; Li et al., 2013; Liang et al., 2020; Ntalli et al., 2010a, 2020; Oka et al., 2000).
The efficacy of these EOs against PPNs has been clearly demonstrated over the past two decades. Oka et al. (2000) showed that EOs of Mentha rotundifolia (isomers of 1,2-epoxymenthyl acetate and piperitone), M. spicata [(−)-carvone and limonene], Origanum vulgare (carvacrol), O. syriacum (carvacrol and thymol), and Coridothymus capitatus (carvacrol) were highly effective against the RKN, causing immobilization of J2s, hatching and root-gall formation inhibition. Carvacrol, thymol and (−)-carvone were particularly noted to be very effective at concentrations >125 μl/l in the in vitro studies; and when mixed in sandy soils at rates of 75–150 mg/kg, root-galling was significantly reduced, with no galls observed on tomato roots raised in pots administered with carvacrol. Similarly, the EOs of O. vulgare, O. dictamnus, M. pulegium, and Melissa officinalis exhibited a high nematicidal activity against M. incognita, with EC50/96h in the range of 1.55–6.15 μl/ml (Ntalli et al., 2010a). At a dose of 250 ppm, Nasiou and Giannakou (2017) showed that carvacrol could paralyze more than 90% of J2s, inhibit egg differentiation and J2 hatching of M. javanica. Laquale et al. (2018) also demonstrated the strong potency of EOs of Monarda didyma and M. fistulosa when tested against M. incognita J2s, with LC50/24h of as low as 1.0 μl/ml. The EOs (eugenol and methyl eugenol) of Agastache rugosa were also found to exhibit significant bioactivity against M. incognita, with a LC50/72h of 47.3 μg/ml (Li et al., 2013); while a relatively higher EC50/72h value of 282 mg/l was revealed when EOs of Ocimum sanctum were tested against M. incognita (Eloh et al., 2020).
Apart from RKNs, several other groups of PPNs have also been targeted using these compounds. At a rate of 2 μg/ml, EOs of Rosmarinus officinalis and Thymus satureioides induced 100% mortality of Xiphinema index within 24 h after treatment (Avato et al., 2017); while a significantly increased mortality of Ditylenchus dipsaci was evident after being exposed to 5,000–7,500 ppm of EOs extracted from O. compactum, O. vulgare, T. vulgaris, and T. mastichina (Zouhar et al., 2009). Additionally, when tested against the pine wood nematode, Bursaphelenchus xylophilus, Barbosa et al. (2010) ascertained that EOs of O. vulgare, Satureja montana, T. capitata, and T. caespititius yielded a minimum inhibitory effect at concentrations of 0.097–0.374 mg/ml, with LC100/24h of 0.858–1.984 mg/ml. Liang et al. (2020) also reported that EOs from Elsholtzia fruticose (mainly eucalyptol, γ-terpinene ã-terpinene and limonene) exhibited a strong suppression effect against D. destructor with a EC50/24h range of 0.16–0.46 mg/ml.
Direct application of water extracts from plant species within the family without the use of toxic solvents has also been tested against PPNs. Water extracts of Mentha spp. known to be rich in flavonoids and carboxylic acids exhibited strong bioactivity against M. incognita, with EC50/72h in the range of 300–1,005 mg/l (Caboni et al., 2013), while water extracts of O. vulgare were highly potent against both J2s of M. incognita and M. javanica, with EC50/96h reported as 27.6 and 37.8 μl/ml, respectively. Additionally, amending O. vulgare powder in nematode-infested soil reduced nematode populations and root-gall formation at an application rate of 5 g/kg soil. A more significant reduction was recorded at a higher application rate of 10 g/kg (Ntalli et al., 2020).
The potency of Myrtaceae
Several species of Myrtaceae are known for the characteristic feature of harboring significant concentrations of foliar EOs (Keszei et al., 2008). Many EOs from these species do not only play the natural defensive roles against pathogens, but are a resource in cosmetic and pharmaceutical industries (Keszei et al., 2008; Rodrigues and Faria, 2021). The genus Eugenia is one of the most important genera within the family. Secondary metabolites (EOs) from barks, leaves, fruits and seeds of ca 350 Eugenia species are known to possess diverse bioactivities against fungi, bacteria and protozoan pathogens (de Souza et al., 2018). The organic extracts of E. winzerlingii exhibit a strong nematicidal effect against M. incognita (Cristóbal-Alejo et al., 2006). Hexane and ethyl acetate extracts of E. winzerlingii are known to be rich in a mixture of fatty acids (decanoic, undecanoic, and dodecanoic acids) which have strong antifeedant and nematicidal properties against RKNs. In particular, Undecanoic acid has been shown to exhibit a significant potency against M. incognita and M. javanica, with EC50/72h values of 192 and 64 nmol/cm2, respectively (Cruz-Estrada et al., 2019). Also, Zouhar et al. (2009) demonstrated that EOs of E. caryophyllata possessed nematicidal activity against D. dipsaci, with a significant mortality rate of up to 93% recorded after a 6-h exposure to a 5,000 ppm solution.
On the other hand, Eucalyptus is also known to contain high foliar terpene-dominated EOs which have been implicated in defensive roles against pathogens (Elbadri et al., 2008; Rodrigues and Faria, 2021). However, Eucalyptus spp. are also known to exhibit qualitative and quantitative variations in EO chemotype composition even among trees of the same species. The monoterpenic aldehyde citronellal, eucalyptol and the ketone piperitone from E. citriodora and E. globulus have been strongly implicated in having significant bioactivity against PPNs (Laquale et al., 2015; Pandey et al., 2000; Rodrigues and Faria, 2021). Other terpenes contained in these two species are: α-pinene, β-pinene, eucalyptol, isopulegol, citronellol, citronellyl formate, β-caryophyllene, β-myrcene, α-terpinene, p-cymene, γ-terpinene, terpinen-4-ol-acetate and α-terpinyl-acetate (D’Addabbo et al., 2020; Rodrigues and Faria, 2021). EOs of E. citriodora showed strong bioactivity against M. incognita, causing almost 100% mortality on exposure of J2s to a 12.5 μg/ml preparation for 8 h; and when applied in soil at a rate of 200 μg/kg soil, significantly lower numbers of eggs and J2s were recovered from tomato roots (D’Addabbo et al., 2020). In addition, Laquale et al. (2015) showed that EOs of E. globulus (rich in eucalyptol) and E. citriodora (rich in the aliphatic terpene aldehyde citronellal) significantly reduced nematode reproduction and root-galling on tomato roots at application rates greater than 50 μl/kg soil. Also, EOs from E. meliodora were found to contain the aromatic aldehyde benzaldehyde, which exhibits a strong nematicidal activity (EC50/24h = 9 μg/ml) (Ntalli et al., 2011). In the Cluster analysis performed by Rodrigues and Faria (2021), EOs of E. globulus were deemed to be quite similar to EOs of other species (E. camaldulensis, E. saligna, E. botryoides, E. polyanthemos, E. viminalis, E. cordieri, E. bosistoana, E. smithii, E. cinerea, and two unidentified Eucalyptus species). EOs of E. camaldulensis, with eucalyptol and α-pinene as the main components have already been tested and implicated in inhibiting egg hatchability, and causing mortality of J2s of M. incognita (El-Baha et al., 2017). This signifies that EOs of many yet to be tested species within the genus potentially have active bioactivity attributes against PPNs.
Compounds from Syzygium spp. have also been evaluated on PPNs. For example, EOs from S. aromaticum, found to be rich in eugenol and β-caryophyllene, exhibited a 95% nematicidal effect on J2s of M. graminicola at the exposure rate of 500 ppm for 72 h (Ajith et al., 2020). Similarly, infestation of M. incognita on tomato was shown to be in check when treated with EOs (eugenol and β-caryophyllene) from S. aromaticum (D’Addabbo et al., 2020). Amending soil with 50 g/kg soil bark powder from S. cumini improved Ricinus communis growth and suppressed M. incognita reproduction and root-galling by 50% (Parveen and Bhat, 2011). Species from other genera like Melaleuca alternifolia are also rich in monoterpene hydrocarbons such as γ-terpinene, α-terpinene, p-cymene and α-pinene, plus oxygenated monoterpenes like β-terpineol, 1,8-cineole and p-menth-8-en-2-ol. These have been found to be highly effective against M. incognita with a LC50/72h of 76.28 μg/ml (Kundu et al., 2020).
The potency of Fabaceae
The family Fabaceae contains species that produce allelopathic compounds that are active in nematode suppression. Such species have been widely used as rotation crops and cover crops (Kokalis-Burelle and Rodríguez-Kábana, 2006). Crotalaria species have widely been utilized as pre-plant cover crops, intercrops, poor or non-host crop or as soil amendments. They are known to produce potentially nematicidal allelopathic compounds like monocrotaline and pyrrolizidine alkaloids (Wang et al., 2002). Additionally, Crotalaria species enhance the microbial activity of some nematode-antagonistic microbiota when incorporated into the soil, including those involved in nutrient cycling (Wang et al., 2002, 2003). For example, incorporation of C. juncea seedlings in cowpea or soybean field significantly reduced M. incognita populations and increased grain yield (Adekunle, 2011). In a rotation scheme performed by Wang et al. (2007), C. juncea and Mucuna deeringiana (velvet bean) effectively suppressed M. incognita, and their effect on reducing nematode infestation was still evident in the subsequent cultivated crop. Velvet bean is known to produce L-3,4-dihydroxyphenylalanine, a compound known to have insecticidal properties (Fujii et al., 1991). Like Crotalaria species, Velvet bean increases the populations of beneficial and nematode antagonistic rhizospheric microbes like Paecilomyces and Burkholderia (Vargas-Ayela and Rodriguez-Kabana, 2001).
Several other species within the family have been directly tested and utilized in the management of PPNs. Extracts from Bauhinia variegate induced significant J2 hatching inhibition and mortality of M. javanica J2s, with LC50/96h of 24.1 mg/ml (Shakya and Yadav, 2020). Piptadenia viridiflora EOs rich in benzaldehyde (98%) were significantly more toxic to M. incognita than the renown carbofuran, causing mortalities of 75–97% at application rates of 100–200 μg/ml, after 48 h. Significant reductions of 90–100% in the number of galls and eggs in tomato roots were also evident when J2s were pre-exposed to benzaldehyde prior to inoculation (Barros et al., 2019a). Also, a 100% nematicidal activity was achieved with EOs from Chamaespartium tridentatum, when applied against B. xylophilus at a rate of 2 mg/ml in a 24-h period. It was found that C. tridentatum contained an assortment of volatiles composed of non-aromatic alcohols, saturated and unsaturated non-aromatic aldehydes, fatty acids and hydrocarbons, with a possibility of a synergistic action against the target organism (Barbosa et al., 2010).
The bark powder from Saraca asoca and Acacia nilotica was shown to suppress M. incognita and Rotylenchulus reniformis reproduction, and subsequently improve castor seedling growth when amended in soil at a rate of 50 g/kg soil (Parveen and Bhat, 2011). In field trials, crude extracts of Tephrosia purpurea were shown to significantly reduce root-galling in French bean (Njenga et al., 2019); while extracts from T. cinerea stem were bioactive against M. incognita (Cristóbal-Alejo et al., 2006). Additionally, extracts of roots and leaves of Leucaena leucocephala have been reported to contain flavonoids like quercetin known to be highly toxic to M. incognita (Adekunle and Aderogba, 2008); Medicago sativa is also known to contain saponin mixture, with nematicidal potency reported against X. index, M. incognita, and G. rostochiensis; and when amended in soil, significant population reductions were evident (D’Addabbo et al., 2011).
The potency of Quillajaceae
Within Quillajaceae, Quillaja saponaria is a species with overwhelming potential as a source of compounds with nematicidal activity, and several commercial botanical nematicide products derived from its extracts have been registered (Table 1). It is an indigenous species to Chile, and is one of the most prime botanical sources of industrial saponins (San Martin and Briones, 1999). Its bark is a ready source of a complex composition of triterpenoid saponins which have been widely used as surfactants, adjuvants, foaming agents and additives in cosmetics and food industries (San Martin and Briones, 1999). However, the triterpene glycoside saponins and polyphenols from Q. saponaria possess fungicidal and nematicidal properties (Chitwood, 2002; San Martin and Magunacelaya, 2005). Whole extracts rich in saponins and other compounds have been shown to be more effective in nematode control, with 100% mortality obtained with 100 ppm treatment after 48–72 h. In field applications at rates of 30 l/ha, similar control efficacy to that of ethoprophos and fenamiphos was achieved (San Martin and Magunacelaya, 2005). Quillaja saponaria extracts were also shown to significantly increase tomato yield in addition to reduction of nematode populations at rates of 15–31 l/ha in greenhouse (D’Addabbo et al., 2006). Similarly, Q. saponaria extract product (QL Agri 35) was effective in controlling RKN in the field, with its efficacy equated to that of cadusafos and oxamyl (Giannakou, 2011).
The potency of Rutaceae
The Rutaceae, like many other families rich in EOs, has its members propertied with bioactive EOs which include limonoids, flavonoids, quinoline alkaloids, coumarins, sesquiterpenes, steroids and triterpenes (Laquale et al., 2015; Lv et al., 2015). EOs and extracts from Evodia and Ruta species are known to possess nematicidal/antihelmintic properties (Ferris and Zheng, 1999; Laquale et al., 2015). Biofumigation with EOs of R. graveolens (main constituents: 2-undecanone and carvacrol) was shown to significantly reduce M. incognita multiplication and gall formation on tomato roots at application rates greater than 50 μl/kg soil (Laquale et al., 2015). Similar results were demonstrated by D’Addabbo et al. (2020) when M. incognita J2s were exposed to 12 μg/ml solution of EOs of R. graveolens. Hatching was moderately inhibited, in addition to a 90% mortality of J2s recorded after 24 h of exposure. Faria et al. (2016) also reported that EOs of R. graveolens showed inhibitory bioactivity against M. chitwoodi population on Solanum tuberosum. On the other hand, Zanthoxylum alatum EOs are rich in Linalool, Limonene and methyl trans-cinnamate; and were found to have a strong nematicidal activity against B. xylophilus, causing 100% mortality at concentration of 5 mg/ml (Kim et al., 2011). Additionally, amending soil with leaves of Z. alatum at a rate of 20 g/kg soil significantly reduced M. incognita multiplication, root-galling; and enhanced cucumber plant growth (Kayani et al., 2012). Among Evodia species, extracts from E. rutaecarpa were shown to be highly effective against M. javanica and Pratylenchus vulnus, causing 100% mortality after 24 h (Ferris and Zheng, 1999).
The potency of Amaryllidaceae (Alliaceae)
Members of the genus Allium are mostly utilized as vegetables and spices. Moreover, many species within the family possess antimicrobial and medicinal properties. Allium sativum (garlic) extract and its constituent EOs is known to exhibit strong antioxidant, antibacterial, nematicidal and antifungal activities (Block, 2010; Jardim et al., 2020; Park et al., 2005; Tibugari et al., 2012). Garlic EOs stand out as one of the most promising botanical nematicides, and when well-prepared, its efficacy is even comparatively better than the renown synthetic carbofuran and dazomet chemicals (Jardim et al., 2020). Its EOs are rich in volatile and non-volatile nematode toxic sulphur compounds, especially the organosulfur allicin, which in itself breaks down to highly toxic sulfur compounds such as diallyl sulfide, diallyl disulfide, diallyl trisulfide, ajoene, and dithiines, among others (Eder et al., 2021; Jardim et al., 2020). Allicin is touted to be the principle active nematicidal compound in garlic. Gupta and Sharmaj (1993) reported that allicin strongly inhibits nematode hatching, in addition to its toxicity. Significantly low hatching rates were observed even at low test rates of 0.5 ppm. In addition, a 100% mortality of M. incognita J2s was recorded at 5.0 ppm within 72 h and a root-dip treatment of tomato seedlings in a 25 ppm preparation for 5 min was found to be effective against M. incognita infestation.
In the recently published studies, Jardim et al. (2020) demonstrated that diallyl disulfide and diallyl trisulfide when isolated and independently tested against M. incognita, were more toxic than carbofuran, with LC50/48h of 134.4 and 36.2 μg/ml, respectively, compared to 151.6 μg/ml for carbofuran. A combination of diallyl disulfide plus diallyl trisulfide yielded even more remarkable results compared to carbofuran and the general EO preparation. Additionally, the vapor of the oil was also shown to be as active as the fumigant nematicide dazomet in the in vitro studies against M. incognita eggs and J2s. Their results were comparable to the findings of Park et al. (2005) who tested purified diallyl disulfide and diallyl trisulfide against B. xylophilus and demonstrated an even lower LC50/4h of 37.06 and 2.79 μl/l, respectively. These findings are comparatively more significant than the findings from tests performed using the commercial NEMguard DE (Certis) (allicin extracted from crushed garlic, with polysulfides 3, 4, and 5 as the main active compounds) against M. incognita (Eder et al., 2021). Most importantly, soil/field treatments show a sustainable protective effect against the target nematode by significantly reducing root-galling and nematode reproduction, with no signs of phytotoxicity (El-Saedy et al., 2014; Jardim et al., 2020; Ladurner et al., 2014). Garlic EOs are therefore potential potent fumigants.
Several other species within the family have also been tested against nematodes. Allium tuberosum extracts (containing carboxylic acids, glycosides, ketones and organic sulfides) were nematicidal against M. incognita and significantly reduced the root-galling on tomato and cucumber (Huang et al., 2016) and extracts of Tulbaghia violacea were effective at reducing M. incognita populations in infested soils (Malungane, 2014; McGaw et al., 2000).
The potency of Euphorbiaceae
The castor bean (Ricinus communis) is the key species of importance within Euphorbiaceae. It is a fast-growing shrub and a highly prolific producer of toxic seeds. The castor bean is utilized as a rotational crop, soil amendment material, compost residues or oil cakes, offering benefits including the release of by-products of decomposition that have nematicidal and suppressive potential against nematodes (Rodriguez-Kabana et al., 1987). When used as rotational crop, castor bean produces and releases allelopathic compounds that are nematicidal or nemastatic as demonstrated by Rodríguez-Kábana et al. (1991). Also, when applied as soil amendments, its residues release organic acids, nitrogen compounds, and other compounds that have potentially adverse effects on nematodes, in addition to the nutritional benefits to plants. The organic matter improves soil structure and stimulates nematode antagonistic fungi and other beneficial microbes. Ammonia is one such by-product of decomposition that is known to be lethal to RKNs (Oka and Yermiyahu, 2002; Rodriguez-Kabana et al., 1987). Specifically, the lectin ricin produced by R. communis is toxic to M. incognita with a EC50 of 33 mg/l for mobility effects (Rich et al., 1989). The oil cake suppresses PPNs when incorporated in the soil (Akhtar and Mahmood, 1996). Soil amendment with leaves of R. communis against M. incognita on tomatoes at rates 15 g/pot was found to promote an increase in tomato plant biomass in addition to significant reduction of nematode populations in the soil and root-galling in plant roots by 80% (Dura et al., 2018).
Other species within the family are also known to produce compounds with nematicidal properties. Extracts from leaves of Jatropha curcas significantly inhibited hatching and caused mortality of M. javanica J2s (LC50/96h = 15.62 mg/ml) (Shakya and Yadav, 2020). Cassava extracts contain methyl palmitate and methyl stearate which are bioactive compounds, and when tested against M. incognita, J2 hatching, egg mass number and root-galling on Musa spp. were significantly reduced (Lu et al., 2020). Also, amending soil with Euphorbia ingens consistently suppressed M. incognita populations in tomato roots, with the suppression effect comparable to that of fenamiphos (Khosa et al., 2020).
The potency of Solanaceae
Solanaceae are a source of important allelopathic secondary metabolites like flavonoids, alkaloids and saponins that are bioactive with a broad spectrum (Eltayeb et al., 1997; Pestana et al., 2010). Solanum spp. are specifically known to contain a variety of steroidal glycosides, oligoglycosides, alkaloids, triterpenoids and flavonoids; compounds that have been investigated for not only their therapeutic properties but also nemastatic and nematicidal potentials (Cai et al., 2010; Pestana et al., 2010). Species like S. sisymbriifolium and S. nigrum when incorporated into soil, control P. goodeyi on banana. They are known to release exudates that have nematicidal effect, in addition to promoting the increase of nematode antagonists in the rhizospheric soils (Pestana et al., 2009).
Solanum scabrum is resistant to both root-knot and cyst nematodes. Using it as rotational crop, Chitambo (2019) demonstrated that soil infestation by root-knot and cyst nematodes was decreased by 85% and 80%, respectively. The suppressive effect was even evident when susceptible cultivars were cropped after 3 seasons of successive cultivation of S. scabrum. Additionally, extracts from species like S. scabrum and other species are toxic to PPNs. For example, in vitro studies showed that extracts of S. scabrum were nematicidal to P. goodeyi; while the acetone extract from S. nigrum caused 100% mortality after 23 h of exposure to a 10 mg/ml extract concentration (Pestana et al., 2010). Standard extracts of various species of Solanaceae (S. nigrum, S. incanum, S. indicum, Hyoscyamus albus, H. muticus, Datura stramonium, and D. arborea) were shown to effectively inhibit hatching of M. incognita in addition to the excellent (100%) nematicidal activity against J2s, 96 h after exposure (Haseeb and Butool, 1996). Datura spp. are also known to contain a variety of bioactive compounds in form of alkaloids, flavonoids, terpenoids, triterpenes, tannins, and saponins. Extracts from leaves of D. metel, D. innoxia, and Brugmansia suaveolens have nematicidal potential against M. incognita (Nandakumar et al., 2017). Hyoscyamus niger extracts at concentrations of 6 and 12% reduced reproduction, inhibited hatching and caused 100% J2 mortality of M. javanica in the in vitro studies (Kepenekçi et al., 2016).
The potency of Lauraceae
Cinnamomum represents an important genus within Lauraceae in terms of its potential in traditional medicine and, as a source of allelopathic secondary metabolites with antifungal and nematicidal properties (Nguyen et al., 2009, 2012). Nematicidal activity of EOs from several Cinnamomum spp. have been documented on several nematode species. For example, EOs of C. verum contain phenolic compounds like E-cinnamaldehyde, eugenol and Z-cinnamaldehyde, and were demonstrated to be highly toxic to M. incognita J2s and eggs compared to the synthetic nematicide oxamyl; causing a 64% mortality at a rate of 0.74 μg/ml after a 24-h exposure in vitro (D’Addabbo et al., 2020). A similar trend was exhibited by C. camphora EOs despite the higher application rate (25 μg/ml) required to attain similar mortality levels. The difference in activity was attributed to the extreme differences in EO composition (C. camphora EOs mainly dominated by terpenes: limonene, eucalyptol and o-cymene). EOs of C. cassia was also found to be rich in E-cinnamaldehyde, and when applied in soil at a rate of 416 μg/ml, root-galling and egg masses of M. incognita in soybean were significantly reduced to levels equivalent to those exhibited by carbofuran applied at similar rates. Additionally, E-cinnamaldehyde has been shown to possess fumigant properties; reducing nematode populations at rates comparable to those obtained with the fumigant dazomet (Jardim et al., 2020). Cinnamomum aromaticum crude extracts were also shown to effectively inhibit infection of M. incognita on cucumber plants (Nguyen et al., 2012); while extracts of cinnamon bark mixed with chitosan were found to effectively inhibit M. incognita and P. coffeae infections on robusta coffee plants under field conditions (Nguyen et al., 2020). Cinnamomum zeylanicum bark and leaf oil (containing mostly cinnamaldehyde and eugenol) was also very effective against M. graminicola, and significantly reduced root-galling in rice (Amarasinghe et al., 2011).
Conclusively, over the course of the past 3 decades, extracts and compounds from a diversity of plant species have been tested against PPNs. The current review cannot exhaust the details of all the studied species. However, the potency of some other notable species from various other families is summarized in Table 2.

A list of examples of plant species and active compounds tested for nematicidal activity against root-knot, cyst and pine wood nematodes
Future Prospects
The global nematicide market is expected to continue growing; and with the increasing call to limit or phase out the use of the most toxic synthetic nematicides, botanical nematicides are without doubt taking a lead as replacements. There is more demand for synthetic chemical-free organic foods and the public has a positive opinion on botanical pesticides (Caboni and Ntalli, 2014). However, there are still unresolved difficulties associated with standardizing the formulations to best utilize the identified natural active constituents without exerting selection pressure on non-target soil microbial communities. Variabilities in quality, efficacy and composition continue to persist due to inconsistencies involved in harvesting, extraction and product formulation methods (Djouahri et al., 2013; Miresmailli and Isman, 2014; Rohloff et al., 2005). Also, despite the overwhelming potential of the compounds extracted from hundreds of plant species, commercialization of new botanical nematicides is generally still lagging, and a large volume of scientific findings remain unutilized due to not only variations in efficacy, but also possibilities of faster breakdown due to the chemical (quick degradation and high volatility) and physical features (quick dispersion) of the identified active compounds. Compounds like benzaldehyde are highly effective against PPNs. However, being unstable, they do not persist in soils to effect long-term control to the target organisms (Barros et al., 2019a). Active compound standardization therefore seems to be one of the missing links to ensure consistency in efficacy. Crude plant products or semi-refined mixtures seem to provide consistent efficacy. However, application of crude plant materials may only offer a short-term solution in small scale farming, as production for use on industrial scale may not be economically feasible in terms of production costs and space. Therefore, improved novel formulation methods that can preserve the chemical composition analogues and compartmentalization of plant active compounds are still needed to minimize environmental concerns, and improve their performance and persistence in soil.
The degradation of the both volatile and non-volatile organic compounds in soil is influenced by many factors, including soil temperature, texture, moisture content, microbial populations, organic carbon content and environmental conditions (Cheng, 1992; Dungan et al., 2003). In order to improve the efficacy and persistence of these allelopathic compounds, an extensive understanding of the effects of soil microbiology, biochemistry and environmental conditions on the retention and transformation of these active components is necessary. Undoubtedly, there is literature on the effects of some of these factors. For example, Borek et al. (1995) found that the transformation and dissipation of allyl ITC and allylnitrile differed with respect to soil physical conditions. Dahlin and Hallmann (2020) demonstrated the significance of soil type in influencing the performance of allyl ITC when treated against M. hapla. Allyl ITC concentrations >20 μmol/mL completely inhibited nematode development in sand while the same concentration when applied in organic potting substrate or normal soil could hardly reduce cucumber root-galling. The degradation of compounds like Methyl ITC is significantly accelerated by increased temperature and organic amendments in soil (Dungan et al., 2003). Unfortunately, many of the successful tests are carried out under controlled experimental conditions with limited possibilities of manipulating these factors. There is therefore a gap between the efficacy of especially volatile compounds and EOs in the usually controlled conditions and the constantly varying field conditions. It is therefore important that scientists and manufacturers of these purified compounds carefully consider and manipulate the soil and environmental factors during trials to improve the product performance in the field.
Additionally, the performance of the purified compounds often differs according to the target nematode species yet many manufacturers tend to generalize the efficacy of these products in the manufacturer’s product labels to generally cover all common plant-parasitic nematode species. These claims are in most cases not backed up by field trials. Additionally, nematode populations in the field are not always uniform. In that regard, the possibility of producing broad spectrum compound products should be studied. For example, Soler-Serratosa et al. (1996) demonstrated that by using combinations of thymol and benzaldehyde, a significant synergistic suppression effect against M. arenaria and H. glycines populations could be achieved. A combination of organic amendments and benzaldehyde was also shown to be effective against M. incognita and other PPNs. This combination significantly reduced root-galling, and nematode populations in addition to improving crop growth in greenhouse trials (Chavarría-Carvajal et al., 2001; Kokalis-Burelle et al., 2002). Also, Flor-Peregrín et al. (2017) showed that nematode suppression and increase in plant growth was more pronounced when garlic extract was combined with mycorrhizal fungi. However, neem and thyme extract instead showed negative interaction with mycorrhizal fungi. Producers therefore should analyze the compatibility and possibilities of blending and stabilizing extracts or compounds from different sources, including the use of biological control agents to improve the spectrum of the final products.
It is also now evident that some isolated active compounds may or may not function in isolation. Faria et al. (2016) and Ntalli et al. (2020) demonstrated that when some EOs are used in isolation, there are possibilities of the compounds being phytotoxic or detrimental to the non-target soil microbial communities. There is therefore, a need to intrinsically understand the role of each constituent in the semi-refined mixtures in neutralizing the unwanted toxic and detrimental effects on both the plant and soil microbial community. Commercial production of the most sensitive compounds should therefore endeavor to take into consideration the possibilities of maintaining and preserving a blend of different chemical constituents of the extracts based on the roles they play to bring about the final effect on the target organism while inflicting limited negative effects to non-target organisms. And lastly, it is apparent that more advanced extraction methods continue to be developed (Mason et al., 2011; Schneider et al., 2011; Sowbhagya and Chitra, 2010). However, such advances are more progressive in the developed world. Developing countries are lagging behind in terms of chemical identification, extraction and formulation techniques, yet many of the promising plant species acting as sources of the active compounds are indigenous in such countries especially in the tropics. Developers should therefore, explore the possibility of coordinating and partnering with such regions with rich sources of the raw plant materials for sustainable production.
Acknowledgments
This work was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture and Forestry (IPET) through Agriculture, Food and Rural Affairs Convergence Technologies Program for Educating Creative Global Leader Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA) (no. 321001-03).
Notes
Conflicts of Interest
No potential conflict of interest relevant to this article was reported.