

Introduction
Gaeumannomyces graminis var. tritici (Ggt), a soilborne pathogenic fungi of wheat roots, causes “take-all disease” on roots of wheat and other cereals and results in severe crop losses worldwide. Take-all disease is an area of intense current interest of research on wheat root diseases yet remains the most important root disease of wheat worldwide. The genetic improvement of resistance in wheat to this pathogen remains a priority, and the study of pathogenic gene expression patterns is crucial for understanding the molecular mechanisms underlying the infection process, providing valuable information for wheat resistance breeding programs.
According to our previous RNA-Seq analysis (Yang et al., 2015), 3,258 DEGs (2,107 up-regulated genes and 1,151 down-regulated genes) were detected during infection process of Ggt. Hence these candidate genes were presumed to play important role in infection process. However, how these genes regulate infection process is still unclear. The more accurate and precise expression pattern of every candidate gene would be helpful to understand how it plays a crucial part. RT-qPCR has become a powerful and frequently used experimental method to evaluate gene expression levels due to its high sensitivity, specificity, accuracy, and reproducibility (Bustin, 2002; Bustin et al., 2005; Nolan et al., 2006). Furthermore, in some cases, it is the only method that allows for the detection of low copy numbers of genes of interest and slight variations in gene expression (Bustin, 2010). Regardless of its many advantages, this method requires appropriate reference genes to ensure the reliability of the target gene’s expression. Recent studies have shown that no single reference gene is universally applicable for all systems. Those genes (e.g., 18S rRNA, actin, tubulin, et al.) involved in basic cellular processes have been frequently used as reference genes. However, many studies have demonstrated that some of these reference genes might not be stably expressed under some conditions (Gutierrez et al., 2008; Schmittgen et al., 2000). For example, Gapdh, which is typically used as reference gene, shows the lowest expression stability among different Aspergillus niger AB1.13 culture conditions (Bohle et al., 2007). Similarly, the research of Kim and Yun (2011) found that GAPDH is an unsuitable reference gene, while GzUBH and EF1A are most stable reference gene of Fusarium graminearum PH-1 strain under different culture conditions. In rust-infected wheat, ADP-ribosylation factor and RNase L inhibitor-like protein (RLI) encoding genes showed the best stability in Puccinia Striticina-infected wheat, while the cell division control (CDC) protein and RLI encoding genes showed the best stability in Puccinia Striiformis-infected wheat (Scholtz and Visser, 2013). However, in Puccinia graminis f.sp. tritici-infected wheat, CDC, 18S rRNA, and β-Tubulin showed the best stability. Thus, a credible reference gene should be evaluated before performing RT-qPCR studies. Many studies have addressed the selection of reference genes in plants and fungi, but none have done so in Ggt, a harmful fungus. This study was conducted to identify reference genes for transcript normalization of pathogenic genes during the infecting process of Ggt. The expression profiles of nine reference genes including GAPDH (Glyceraldehyde-3-phosphate dehydrogenase), TUB2 (tubulin alpha chain), G6PDH (Glucose-6-phosphate-1-dehydrogenase), GK (Glucokinase), EF-3 (Elongation factor 3), EF2-2 (Elongation factor 2), TUBβ (Tubulin beta), ACT (Actin) and EF2-1 (Elongation factor) were studied in different stages of Ggt-infected wheat, and the expression stability of these genes was evaluated with geNorm (Vandesompele et al., 2002), NormFinder (Andersen et al., 2004) and Bestkeeper (Pfaffl et al., 2004). Furthermore, the expression pattern of target gene ExoPG (exopolygalacturonase) was investigated using the selected reference gene, which may be helpful in revealing its role in Ggt pathogenicity.
Materials and Methods
Fungal meterials and infection of wheat roots
An isolate of Ggt (GGT-007, Henan Academy of Agricultural Sciences, Zhengzhou, China) was isolated by Quan et al. (2014). Liquid cultures were grown in potato dextrose broth at 25°C with shaking at 180 rpm for 5 d to prepare for wheat root infection and Ggt RNA extraction.
Seeds of winter wheat (Aestivumtriticum cv ‘Zhengmai 366’) were supplied by the Wheat Research Institute of the Henna Academy of Agricultural Science. The experiment of accelerating germination and inoculation procedure of wheat root were carried out according to Yang et al. (2015a). The wheat roots infected by Ggt samples from three biological replicates were collected at 0.5, 1, 2, 3, 4, 5, 7 d after inoculation. The liquid culture of Ggt was control group. The samples were immediately frozen and then stored in liquid nitrogen until analysis.
Extraction of total RNA and synthesis of cDNA
Frozen Ggt hypha and infected-wheat roots were ground to a fine powder in liquid nitrogen with a pestle and mortar. About 100 mg of powder was used for RNA extraction. Total RNA was isolated using the RNAiso Plus (TaKaRa, Dalian, China). Purified RNA was quantified with Nano-Drop2000 spectrophotometer (Thermo, Wilmington, DE, USA), and the concentration and integrity of RNA was checked by a denaturing 1.0% (p/v) agarose gel. Only RNA samples with 260/280 wavelength ratio between 1.9 and 2.1 were used for cDNA synthesis. cDNA was synthesized with 1 μg total RNA according the manufacturer of the Prime Script™ RT reagent Kit with gDNA Eraser (Perfect Real Time) (TaKaRa, Dalian, China) in a total volume of 20 μl cDNA was stored at −20°C.
Primer design and PCR conditions
The nine candidate reference genes (PFK, GAPDH, TUB2, G6PDH, GK, EF-3, EF2-2, TUBβ, ACT and EF2-1) and one target gene ExoPG were selected from Transcriptome Sequencing Data and Genebank. Their accession numbers are listed in Table 1. Primer sequences were designed using Primer 3.0 (primer3.ut.ee/) with the following parameters: Tm around 60°C and product size ranges 150-250 base pairs, primer sequences with a length of 18 to 22 nucleotides, and a GC content of 45-55%. All the primers’ specificities were confirmed using Primer-BLAST searches on nr database of NCBI and analyzed by real-time PCR and electrophoresis on 2% agarose gel.
QRT-PCR reactions were performed in 96-well plates with Step One Plus Real-Time PCR System (Applied Biosystems, CA, USA) and a SYBR® Premix Ex Taq™ Kit (TaKaRa, Tokyo, Japan). The qRT-PCR conditions were as follows: 95°C for 30 s and 40 cycles of 95°C for 5 s and 60°C for 30 s. The amplicons specificities were detected by melting curves analysis (60 to 95°C) after 40 PCR cycles.
Analysis of candidate reference gene expression stability
For every primer pair, a standard curve was used to calculate the gene-specific PCR efficiency from the 5-fold series dilution of Ggt cDNA. The correlation coefficients (R2) and slope value were determined from the standard curve. The following formula was used to calculate the qRT-PCR efficiencies (E). E = (5−1/slope − 1) × 100 (Radonić et al., 2004).
The expression stabilities of all the candidate genes were evaluated by three Excel-based softwares: geNorm (Vandesompele et al., 2002), BestKeeper (Andersen et al., 2004), and NormFinder (Pfaffl et al., 2004). The geNorm (http://medgen.ugent.be/~jvdesomp/genorm/) is based on the mean pairwise variation between all the tested genes to calculate the expression stability value (M) of genes, and BestKeeper (http://www.gene-quantification.de/bestkeeper.html) is used to evaluate the expression stability according to an index which is obtained by standard deviation (SD) and percentage covariance (CV). NormFinder (http://www.mdl.dk/publicationsnormfinder.htm) is used to rank the genes expression stability according to their intra- and inter-expression variation.
Results
Identification of reference gene candidates and primer specificity and efficiency
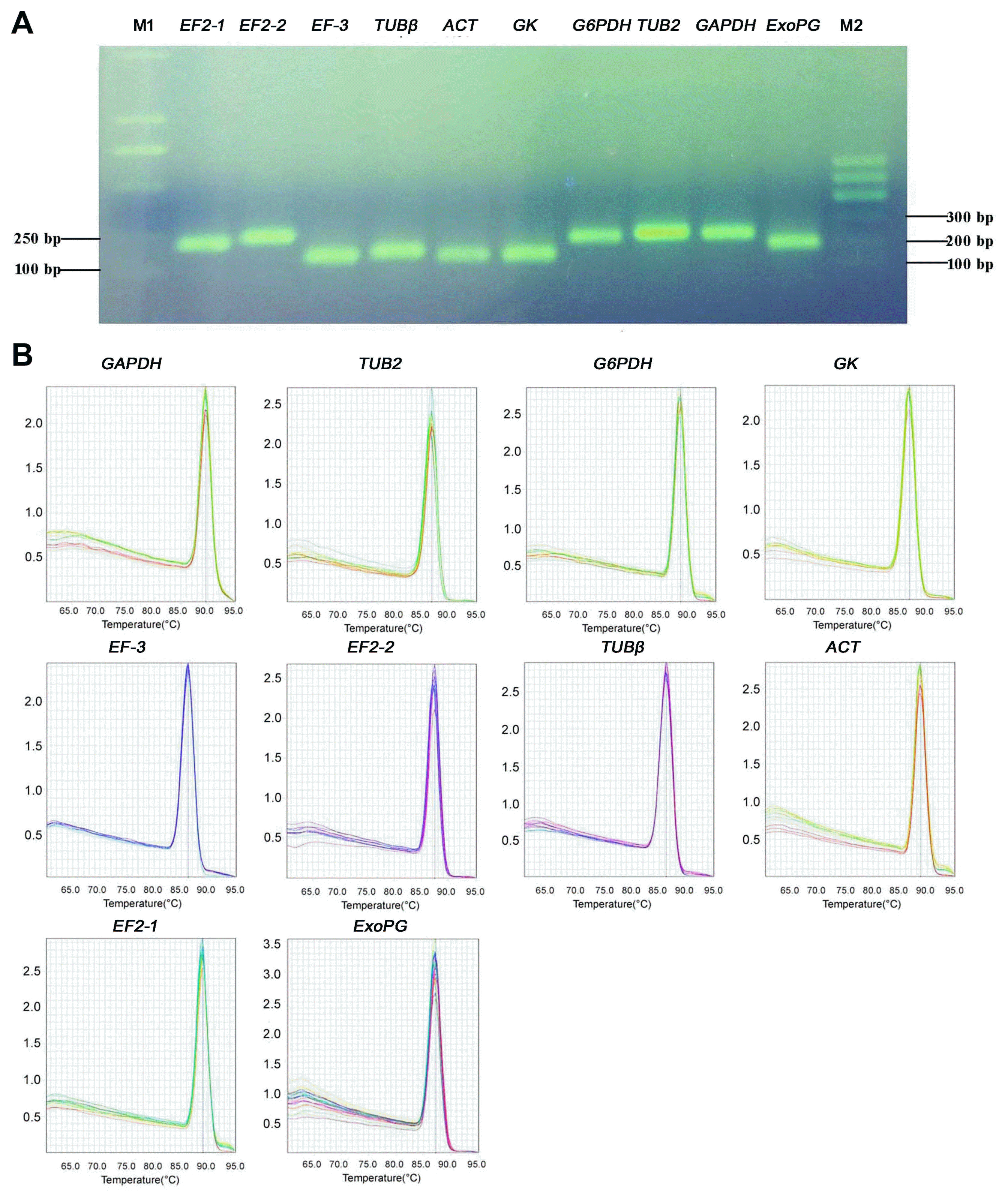
Nine reference gene were selected for RT-qPCR normalization. Gene name, accession number, gene description, primer sequences, melting temperatures value (Tm), amplification efficiency and correlation coefficients are listed in Table 1. The Tm of PCR products ranged from 86.2°C for EF-3 to 89.63°C for GAPDH. Amplification efficiency (E) of PCR reactions ranged from 103.7 for GAPDH to 119.82 for EF-3. Correlation coefficients (R2) ranged from 0.9853 for TUB2 to 0.9999 for EF-3. The amplifications were confirmed by the presence of a single band in 1.5% agarose gel electrophoresis and by the single peak melting curve of the PCR products (Fig. 1A, B), and the size of PCR products were consistent with expected size for each primer pairs.
Expression levels of the selected candidate reference genes
Based on average cycle threshold (Ct) values, there were obvious differences in transcript levels among the reference genes during Ggt infection. The Ct range and coefficient of variance for each gene were also obtained from the Ct value. As expected, the average Ct values varied from 20.56 for GAPDH to 25.77 cycles for GK (Fig. 2). Most of them were expressed abundantly (ranging from 20 to 25). Among them, GAPDH was the most abundantly expressed reference gene, while GK was the least abundant. According to the coefficient of variance (CV), which can reveal the reference gene’s expression stability, EF-3 had the least variation with a CV value of 4.17%, while GAPDH had the most variation with a CV value of 7.43%.
Expression stability of candidate reference genes
Ct data were obtained from qRT-PCR. Ct values were used directly for stability calculations for BestKeeper, while Ct values were converted to relative quantities for geNorm and NormFinder.
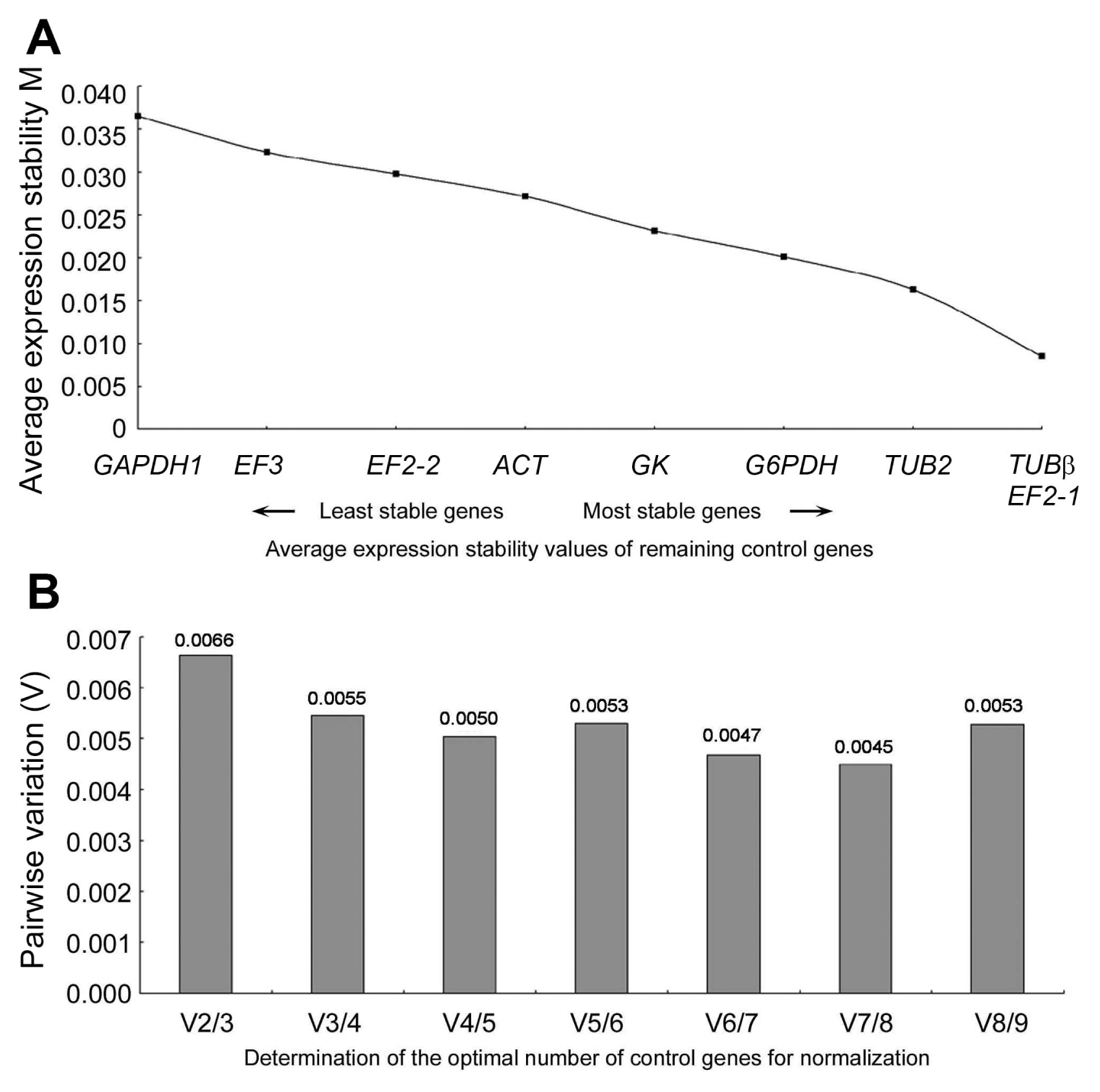
Gene expression stability has a negative correlation with its M value. It was calculated by geNorm software based on the average pairwise variation between all the tested reference genes. The most stable reference gene has the lowest M value. According the Fig. 3A, all the reference genes can be recognized as stable because they all had an M value less than the geNorm threshold of 1.5. During the process of pathogen infections of wheat roots, TUBβ and EF-2 had the highest expression stability (i.e., the lowest M values, 0.028 and 0.03), whereas GAPDH had the lowest stability (M value = 0.051). The other six genes were in between.
To select the optimal number of reference genes for credible normalization, pairwise variation (Vn/Vn+1) between two sequential normalization factors NFn and NFn+1 was calculated by geNorm (Fig. 3B). The result showed all pairwise variation to be smaller than the recommended threshold (0.15). Therefore, one reference genes would be sufficient for proper gene expression normalization throughout the infecting period.
NormFinder, an Excel-based software tool, evaluates the most stable reference genes according to the stability value based on their intra- and inter-expression variation. Those with lower average stability values are regarded as more stable expression genes (Table 2). TUBβ was regarded as the most stable reference gene.
BestKeeper is used to rank the candidate genes based on their coefficient of correlation (r) relative to the BestKeeper index, by calculating the Ct set standard deviation (SD) and coefficient of variance (CV). The results (Table 3) revealed the best correlations were obtained for TUBβ (0.991) with the P value of 0.001, while the lowest correlations were obtained for EF3 (0.891), with a P value of 0.003. Therefore, TUBβ was ranked as the most stable reference gene, which was in accordance with the results obtained from geNorm and NormFinder. In general, the results of Best Keeper analysis showed few differences from those obtained from geNorm and Normfinder.
Validation of the selected reference gene
To verify the reliability of the selected reference gene, we analyzed the expression pattern of one different expression transcript GGTG_09200 obtained from transcriptome sequencing. Transcript GGTG_09200 code ExoPG belongs to a group of plant cell wall degrading enzymes (PCWDEs), which can damage plant tissue, cell walls and cause leaks of cytoplasm (Kang et al., 2000; Valette-Collet et al., 2003).
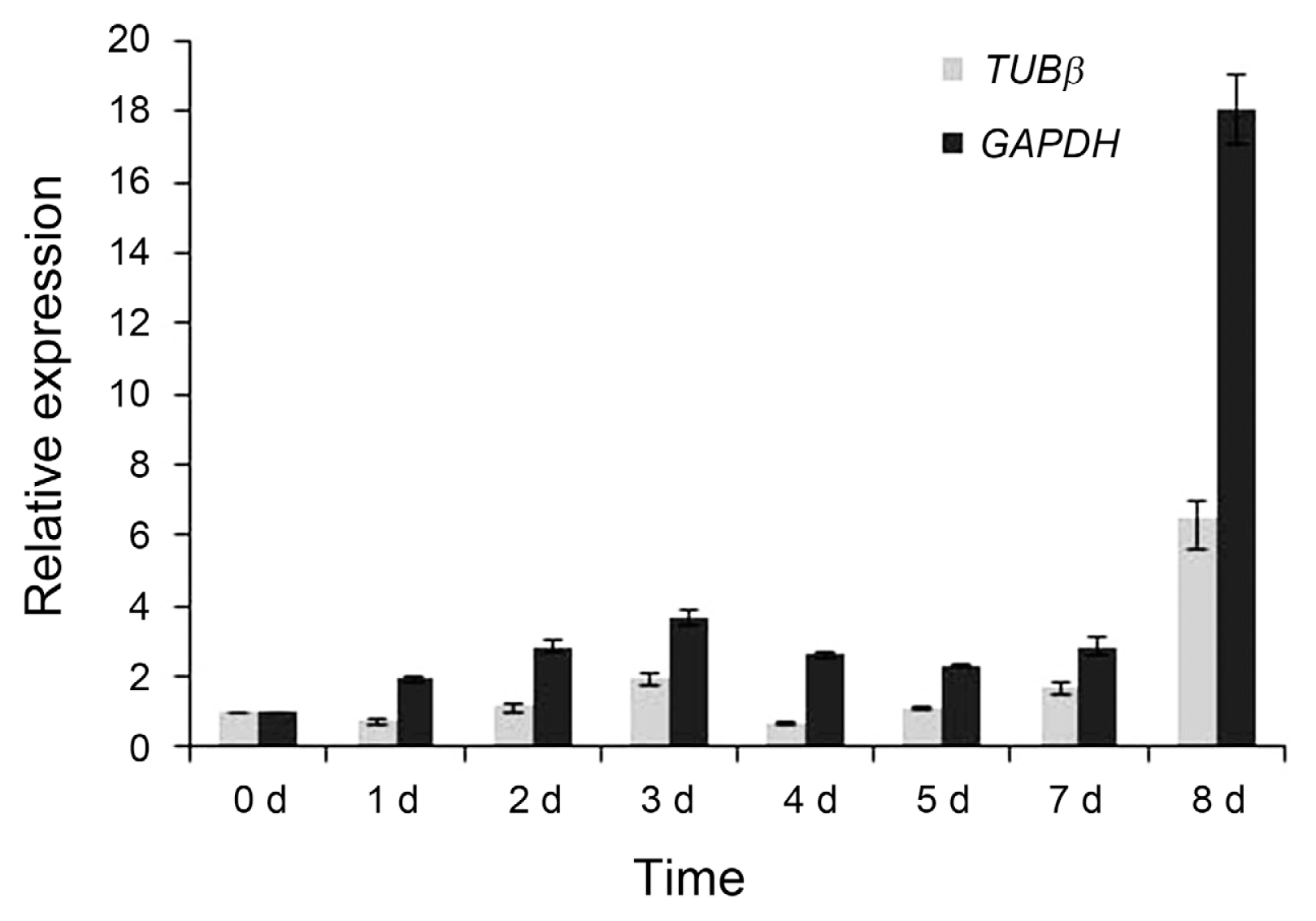
The most appropriate reference gene (TUBβ) and the most inappropriate reference gene (GAPDH) were selected for qRT-PCR normalization of ExoPG, respectively. ExoPG is important in the infection process of plant pathogenic fungi, and it might be activated to degrade pectin and hemicellulose to obtain nutrients or to weaken the wall to allow hyphal growth in the root. The normalization of qRT-PCR using TUBβ generated similar expression patterns with a low expression in earlier stages of infection process and the increase seven days after infection (Fig. 4), which agrees with the results of transcriptome sequencing. When the least stable reference gene GAPDH was used for normalization, the ExoPG gene was expressed at high levels at the beginning of infection.
Discussion
At present, qRT-PCR has significantly improved the detection and quantification of expression profiles of genes of interest due to its high throughput, specificity, and accuracy (Bustin, 2002; Nolan and Bustin, 2006). However, it is important to select appropriate reference genes for gene expression normalization because of the relative expression of target genes can be influenced obviously by reference genes selected. Some commonly used reference genes have come under scrutiny or were doubted in recent studies for their variable expressions when tested with different experimental treatments (Ana et al., 2011; Fang et al., 2006; Hacquard et al., 2011; Huang et al., 2012; Teste et al., 2009; Yan and Liu, 2006).
Root and crown rot of wheat, also called “Take-all”, and caused by fungus Ggt, affects the quality and yield of wheat and other Gramineae crops. We might effectively improve the disease resistance of wheat through genetic improvement by understanding its intrinsic characteristic essentially of pathogenic (rewrite whole sentence). While up to now, the pathogenesis is unclear. Taking advantage of transcriptome data (Yang et al., 2015), a precious sequence resource for non-model fungus, and many pathogenicity-related genes will be identified with comprehensive functional analysis in the near future. In this study, we performed a selection and validation of internal reference genes for further accurate gene expression analysis using RT-qPCR in in the infection process of Ggt.
Although some previous studies have successfully used one or two of the three excel-based program geNorm, NormFinder and BestKeeper to select reference genes (Ana et al., 2011; Huang et al., 2012), we suggest that taking into consideration the stability ranking of all three programs would provide a certain degree of confirmation of the results. Although there were differences in the rank of the candidate reference genes according to the three programs, all three analytical programs used in this study found TUBβ to be the most stably expressed reference gene in the infection process of Ggt. ACT is a commonly used housekeeper gene for RT-qPCR expression normalization in fungi, however, it was found to be a relatively unstable reference gene in this study. This finding is consistent with the results of Ana et al. (2011) and Huang et al. (2012). Moreover, TUBβ is regarded as the least stable reference gene during the infection process of Hemileia vastatrix isolate 1065 (Vieira et al., 2011). In contrast, TUBβ was regarded as the most stably expressed reference gene during the infection processes of Ggt in our study. And the results of Yan and Liu (2006) and Huang et al. (2012) all show TUBβ is the most stably expressed housekeeper gene in infection processes of Phytophthora parasitica and Puccinia Striiformis f. sp. tritic, which might provide more accurate expression measurement of target genes.
In the infection progress of fungi, Phytopathogenic fungi expose plant cell walls to cell wall degrading enzymes (CWDEs), CWDEs like ExoPGs are needed for initial penetration and believed to be important pathogenicity factors (D’Ovidio et al., 2004; ten Have et al., 1998; Weste, 1970). To validate the reference gene candidates selected by the three programs, the transcript level of ExoPG gene was asserted during the infection process of Ggt. There was an obvious difference between the relative expression levels normalized by the most stable reference gene and the least one. And the relative expression level of ExoPG gene was consistent with the results of RNA-seq (Yang et al., 2015). These results suggest that the incorrect use of reference genes may introduce bias into the analysis and lead to the misinterpretation of data.
In summary, nine candidate reference genes from Ggt evaluated in the infection process of Ggt using geNorm, NormFinder and BestKeeper programs, the results reveal TUBβ was the most appropriate reference gene for RT-qPCR normalization across the infection process of Ggt. This study constitutes the first systematic exploration in Ggt to identify optimal reference genes for RT-qPCR normalization and useful to understand the functions of candidate target genes.

 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






