

Recently, the use of synthetic fertilizers and pesticides have resulted in deterioration of soil structure and increase in soil-borne phytopathogens (Bailey and Lazarovits, 2003). As a result, along with many phytopathogenic fungi such as B. cinerea, F. oxysporum, P. capsici, R. solani, and S. cepivorum etc. having a wide range of host plants, it has been very difficult to control the spread of the plant disease. Although applying fungicide has been one of the conventional methods for controlling fungal diseases, there is increasing international concern over the use of fungicides on crops because of their harmful effects on human health and the emergence of pathogen resistance to fungicides. To cope with this problem, various disease managements including biological control, have been applied.
Biological control by microorganisms (Huang et al., 2012; Kim et al., 2012; Li et al., 2012) has received considerable attention as a reliable substitute for the use of various hazardous chemical fungicides, which have raised serious concerns of food and environmental contamination (Fernando et al., 2005). Microorganisms also play an essential role in several life support functions as they enable soil to recycle nutrients, suppress plant pathogens (Mendes et al., 2011), and serve as a suitable substrate for plant growth (Kim et al., 2011). Among the myriads of bacteria in the plant rhizosphere, a few spore-forming plant growth promoting rhizobacteria (PGPR) attract special attention due to their stable habitancy in soil (Emmert and Handelsman, 1999; Jin et al., 2006).
Soil samples were collected from the Chinese cabbage, garlic, and paddy fields in 30 regions of Samcheok, Gangwon province, South Korea during November 2011. Each sample was air-dried at ambient temperature and then passed through an 850 μm mesh sieve to remove plant debris. Soil samples were preserved in polyethylene bags at room temperature before use. Soil suspensions were prepared in sterile water and cultured on the surface of potato dextrose agar medium (PDA, Difco, USA) and tryptic soy agar medium (TSA, Difco, USA). The plates were incubated at 25°C for 10 days. Afterwards, colonies with different morphological appearances were chosen from the countable plates and streaked on fresh similar medium to obtain pure cultures.
Bacterium was identified by morphological and taxonomic characterization and 16S rRNA gene sequencing assay. Morphological characteristics were examined on an SEM at an accelerating voltage of 10 kV. Taxonomic characterization of the strain was performed on the basis of test for oxidase, catalase, H2S production, gelatin hydrolysis, starch hydrolysis, nitrate reduction, urase hydrolysis, casein digestion, arginine dihydrolase, ornithine decarboxylase, esculin hydrolysis and utilization of carbon such as glucose, sucrose, dulcitol, salicin, D-mannitol, D-adonitol, L-arabinose, D-xylose, and D-trehalose (Takagi et al., 1993). The results of these tests were scored as either positive or negative.
The total genomic DNA was extracted as described by Nishiguchi et al. (2002). The 16S rRNA gene was amplified by polymerase chain reaction using universal primers 27F and 1492R (Bioneer, Korea). The amplified DNA was purified using a gel purification kit (Bioneer, Korea) and was sequenced at Bioneer. Similarity searches of the sequence obtained was performed using the BLAST function of GenBank.
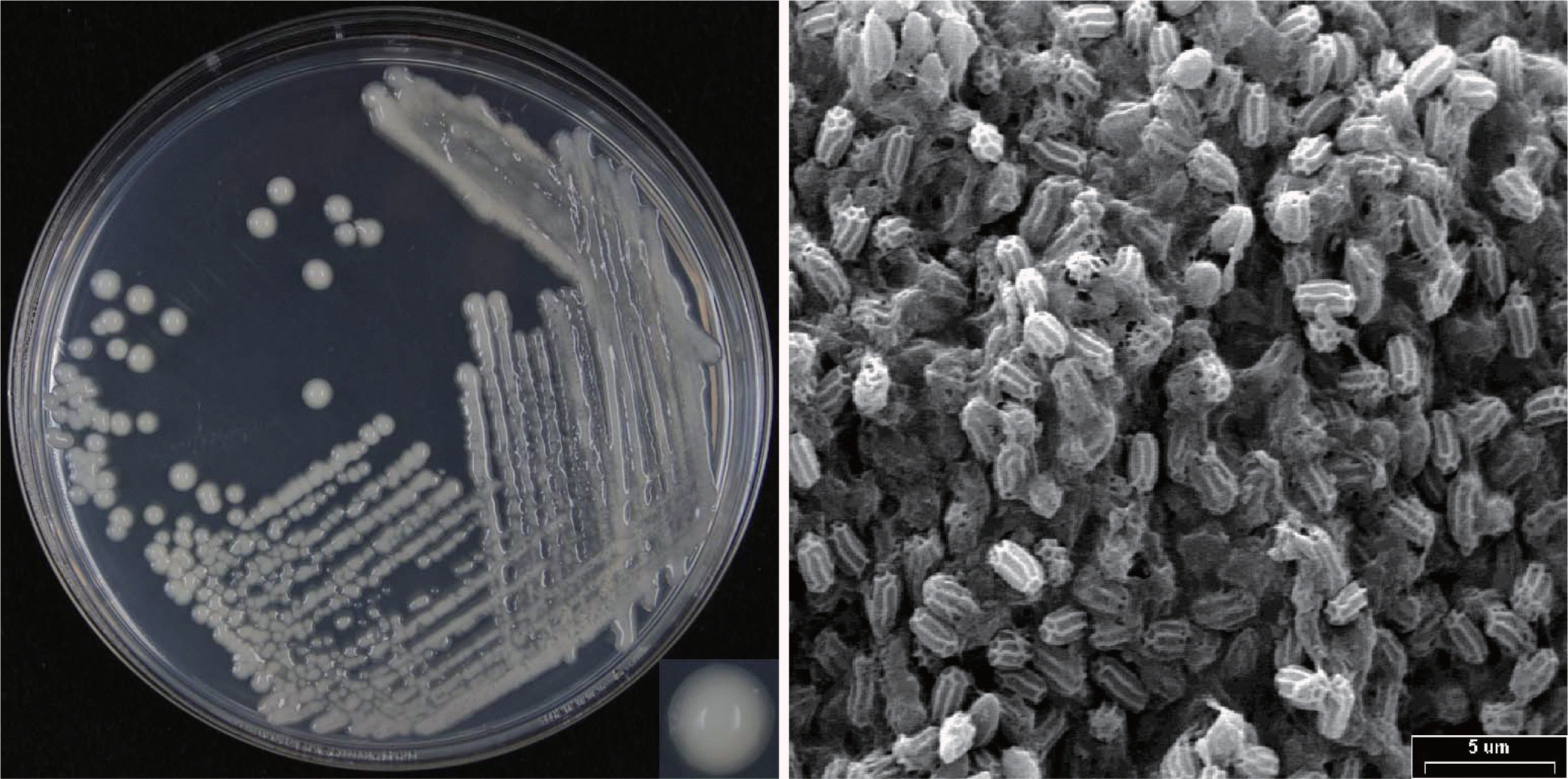
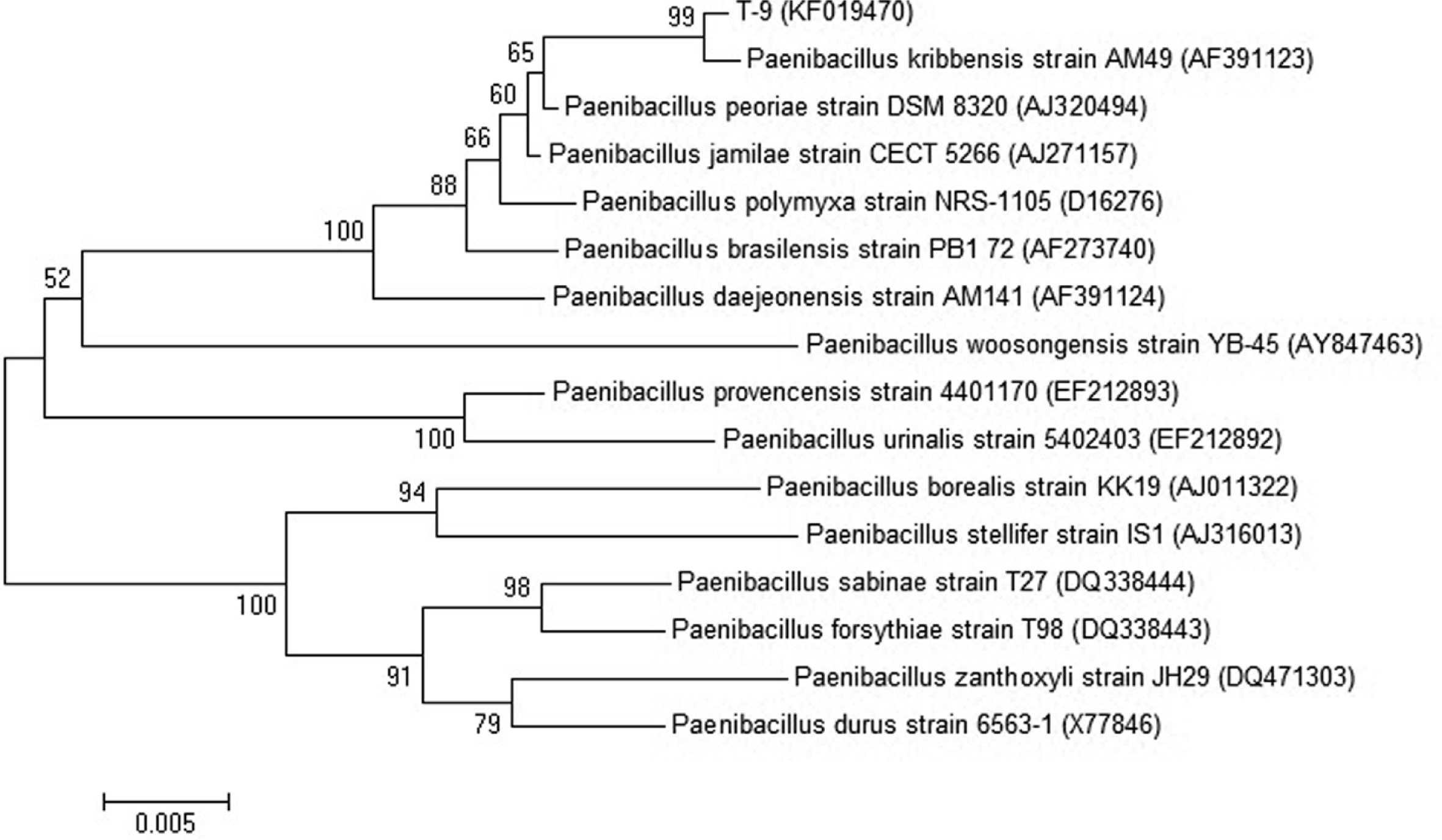
The predominant strain obtained from soil samples of Samcheok was named as T-9 and found to be a Gram-variable, rod-shaped, and facultative anaerobic bacterium. It produced flat colonies and unsmooth-surfaced spores (Fig. 1) and catalase (Table 1) although it did not produce H2S. Methyl red reaction was positive while Voges-Proskauer reaction was negative. An array of carbon sources were utilized like glucose, lactose, sucrose, salicin, L-arabinose, raffinose, maltose, and cellobiose, but not dulcitol, D-adonitol, myo-inositol, and L-rhamnose. The 16S rRNA gene sequence analysis indicated that the closest relative (99%) of the strain T-9 (1465 bp) was P. kribbensis AM49 (AF391123) (Fig. 2). The 16S rRNA gene sequence of strain T-9 was deposited in the GenBank database with an accession number KF019740. From the results of morphologic and biological characteristics, and 16S rRNA gene sequence analysis, the predominant strain T-9 obtained from soil samples was identified as P. kribbensis and thus named P. kribbensis strain T-9.
The genus Paenibacillus, originally included within the Bacillus and reclassified as a separate genus (Ash et al., 1993), is prevalently distributed in different soils and plant rhizospheres (Berge et al., 2002; von der Weid et al., 2002; Garbeva et al., 2003; Yoon et al., 2003). Different species of Paenibacillus have been demonstrated as having potential antifungal activities (Fortes et al., 2008; He et al., 2007; Liu et al., 2008; Raza et al. 2008; von der Weid et al., 2003; von der Weid et al., 2005). The P. kribbensis is recently described as a new Paenibacillus species isolated from soil (Yoon et al., 2003). Some strains produced antimicrobial substances, suggesting an important role in the soil. However, no biological role of this species in the rhizosphere have been reported.
The antifungal activity of P. kribbensis strain T-9 against the phytopathogenic fungi was performed by dual culture. Briefly, a mycelial column (6.5 mm in diameter) was cut off from the white perimeter portion of the colony, which had been cultured on PDA plates at 25°C for 7 days, and placed at a distance 1.5 cm away from the rim of a fresh PDA plate. Afterwards, the P. kribbensis strain T-9 growing in PDA was sowed in a hole with a sterilized toothpick on the PDA plate (1.5 cm away from the other rim). Simultaneously, control plates inoculated in different mycelial columns were prepared without P. kribbensis strain T-9. The antagonistic effect of the control plates was evaluated by measuring the size of the inhibition zone on the PDA plates following culturing at 25°C for 14 days.
The P. kribbensis strain T-9 showed strong antagonistic activity against a wide range of phytopathogenic fungi that cause disease on different crops (Table 2). It produced the highest antifungal activity against P. capsici, which was cosmopolitan in distribution. Furthermore, there was a moderate antifungal activity against B. cinerea, C. acutatum and M. oryzae. Other test phytopathogenic fungi were also inhibited, but the antifungal activity was relatively less.
The in vivo efficiency of P. kribbensis strain T-9 for control of anthracnose in pepper fruits, Phytophthora blight in pepper plant, blast, sheath blight in rice plant and gray mold in tomato plant, was evaluated in plastic sieve systems or under greenhouse conditions (Table 3). Pepper (Capsicum annuum) cv. Super manitta and Hongjinjoo, rice (Oryza sativa) cv. Nakdongbyeo and tomato (Solanum lycopersicum) cv. Super dotaerang were sown in horticultural soil in seedling plate. The plants were kept in the greenhouse for one month. To obtain culture broth, P. kribbensis strain T-9 was cultured in potato dextrose broth medium (PDB, Difco, USA) for three days, which included incubation in a rotary shaker at 200 rpm and 25°C. After incubation, P. kribbensis strain T-9 suspension was diluted in distilled water to 108 cfu/ml (OD620=0.1). Each seedling and fruit was treated with growth broth one day before inoculation with B. cinerea, C. acutatum, M. oryzae, P. capsici, and R. solani. For pepper anthracnose, each fruit was soaked in culture broth, water, or propineb for 30 min. For pepper Phytophthora blight, each seedling was dip-treated with culture broth, water, or dimethomorph for 10 min. Also culture broth, water, or fungicides were sprayed on each seedling for other plant diseases. Chemical fungicides were used as the positive control and water was used as the negative control. The inoculated plants were placed in a moist chamber to maintain approximately 90% humidity at 25°C for 24 h, and then transferred to the growth room for further incubation. There were three replicates for each treatment.
The P. kribbensis strain T-9 for control of Phytophthora blight in pepper plant, blast, sheath blight in rice plant and gray mold in tomato plant, was highly effective, reducing disease severity by 92.9%, 78.5%, 100%, and 89.8% relative to the inoculated control, respectively. The chemical fungicides were used as reference, which reduced disease severity by 99.3%, 74.3%, 100% and 93.2%, respectively. The in vivo efficacy of P. kribbensis strain T-9 against anthracnose in pepper fruits was evaluated in plastic sieve systems. The P. kribbensis strain T-9 was highly effective, which showed a control efficiency of 96.7%. The synthetic fungicide was used as reference, which showed a 92.3% control value.
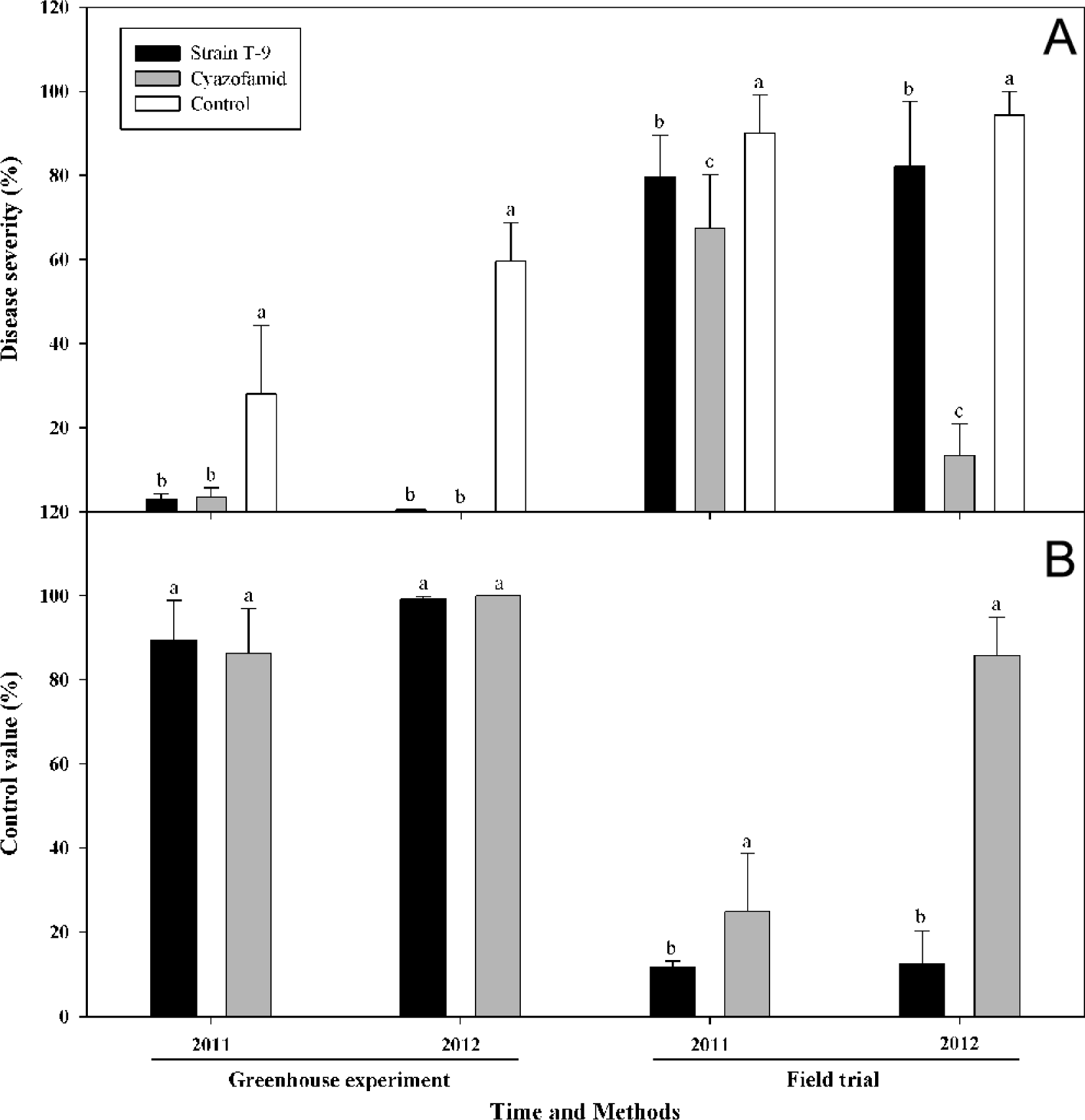
The suppressive effects of P. kribbensis strain T-9 against Plasmodiophora brassicae in the greenhouse and the field were assessed in 2011 and 2012, respectively (Fig. 3). Chinese cabbage (Brassica campestris) cv. Gaeulmas, a highly susceptible to clubroot, was sown in horticultural soil in seedling plate. The plants were kept in the greenhouse for one month. To obtain culture broth, the method was same in upper. Frozen galls were soaked in distilled water for a while to soften the tissue, and then they were macerated in a homogenizer with distilled water at high speed. For inoculation, inoculate suspension was mixed into horticultural soil. In 2011 and 2012, Chinese cabbage was transplanted in a plastic pot containing the clubroot-infested horticultural soil. Each seedling was dipped into culture broth. Cyazofamid was used as the positive control and water as the negative control. The seedlings were watered regularly to keep constant soil moisture during cultivation in the greenhouse. After forty days, each root was rinsed with tap water and assessed for clubroot severity based on a 0-4 scale, where 0 indicated no infection; 1 indicated 1-25% of roots infected; 2 indicated 26-50% of roots infected; 3 indicated 51-75% of roots infected; and 4 indicated more than 75% of roots infected. Statistical analyses were conducted with the Statistical Analysis System for personal computers (Ver. 9.2, SAS Institute Inc.). Fisher's protected least significant difference (LSD) at P ≤ 0.05 was applied to determine whether or not differences between treatments were significant.
In 2011 and 2012, a study using Chinese cabbage was conducted to assess the efficacy of microbes in the field, based on the efficacy observed in a previous experiment. Inoculate suspension was mixed into field soil using a rotary cultivator. Each seedling was dipped in culture broth for 10 min. Seedlings were transplanted in the infected field and cyazofamid was used as the positive control and water as the negative control. The trials were arranged in a randomized complete block design with three replicates. The seedlings were watered regularly to keep constant soil moisture. After seventy days, each root was rinsed with tap water and assessed for clubroot severity based on the 0-4 scale mentioned above.
For greenhouse experiment, the P. kribbensis strain T-9 was highly effective against clubroot, which showed a control efficiency of 89.3% and 99.2% in 2011 and 2012, respectively. Cyazofamid was used as a reference, which showed 86.3% and 100% control values. For field trials, P. kribbensis strain T-9 had no substantial impact on clubroot severity, with 79.6% and 82.2% disease severity on the susceptible cultivar of Chinese cabbage, which reduced the disease severity by 11.7% and 12.6% in 2011 and 2012, respectively. The cyazofamid treatment reduced the disease severity by 24.9% and 85.8%.
The stratagem of controlling the biological properties of the P. brassicae has become an important approach for creating a long-lasting effect on the facilitation of sustainable agriculture; however, only a few antagonistic microorganisms have been reported, including Micromonospora rosea subsp. rosea and Streptomyces spp. (Kim et al., 2004; Lee et al., 2008; Wang et al., 2012), Streptoverticillium olivoreticuli ssp. (Kim et al., 2002), Cellulosimicrobium cellulans, and Paenibacillus spp. (Choi et al., 2007). The progress and applicable fields of biological control for clubroot are limited compared to other soil-borne plant diseases, and there are no commercial biological control products available. The continuous search for antagonistic agents against clubroot is necessary. In this study, the suppressive effects of P. kribbensis strain T-9 against P. brassicae were assessed in the greenhouse and the field in 2011 and 2012. We found the efficacy observed from the field trial to be lower than that of the greenhouse experiment. For the greenhouse experiment, P. kribbensis strain T-9 showed a good control efficiency of 89.3% and 99.2% in 2011 and 2012, respectively. However, for field trials, there was no substantial impact, but the treatment reduced the clubroot on the susceptible cultivar of Chinese cabbage, with disease severity, by 79.6% and 82.2% in 2011 and 2012, respectively. Basic environmental conditions, such as temperature, moisture, and soil physical and chemical characteristics, can greatly affect the interactions among plant, pathogen, and biocontrol agents in various ways, and all of these effects may influence the efficacy of biological control (Larkin and Fravel, 2002). On the other hand, enormous microorganisms exist in the rhizosphere of plants, which may influence the efficacy of biological control. Finally, endospore formation and longer generation times influence them to rapidly colonize roots, which they directly protect against soil-borne pathogens while they adapt their fitness to that of rhizosphere (Ryu et al., 2005).
In summary, this study is the first antagonistic activity assay of P. kribbensis strain T-9 against various soil-borne phytopathogenic fungi. It was found that the P. kribbensis strain T-9 has the potential to offer protection to plants against different soil-borne phytopathogenic fungi, in addition to its antagonistic activity, suggesting that further studies should be considered for using P. kribbensis strain T-9 as a biological control agent and enhancer of control efficiency in field trials.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






