

Geminiviruses are approximately 2.8 kb single-stranded circular plant DNA viruses that can cause serious losses of crop products worldwide (Vanderschuren et al., 2007). They are divided into four genera: Mastrevirus, Curtovirus, Topocuvirus, and Begomovirus. Beet curly top virus (BCTV) and Beet severe curly top virus (BSCTV) are Curtoviruses that infect only phloem-limited dicotyledonous plants and raise symptoms such as stunted growth, leaf curling, accumulation of anthocyanin, vein swelling and hyperplasia of the phloem (Latham et al., 1997). Geminiviruses regulate several signaling pathways involved in cell cycle regulation and host defense for the purpose of viral propagation. Infection with geminiviruses leads to cell cycle reprogramming using RBR. Viral proteins such as AL1 (also known as AC1, C1 and Rep) and C3 (also known as REn) bind to RBR and inhibit it (Desvoyes et al., 2006; Kong et al., 2000); thus, the infected cells express genes associated with the onset of G1, S and early G2 phases, while suppressing those associated with the early G1 and late G2 phases (Ascencio-Ibanez et al., 2008).
Plants have innate antiviral defense systems such as gene silencing, and regulation of salicylic acid biosynthesis and metabolism (Zhang et al., 2011). Geminiviruses also interfere with these defense systems. To facilitate geminivirus replication, C2 interacts with ADK and SAMDC1 and suppresses the plant methyl cycle (Wang et al., 2005; Zhang et al., 2011), and inhibits SnRK1 (Hao et al., 2003). SnRK1-mediated innate antiviral defense was identified by the interaction of geminivirus C2 proteins with SnRK1.2 (Hao et al., 2003), and it was reported that SlSnRK1 reduces geminivirus infection by interacting with and phosphorylating the βC1 protein (Shen et al., 2011). SnRK1 is a key regulator of plant stress and metabolism, and it regulates global transcription (Baena-Gonzalez et al., 2007). Therefore, it was suggested that SnRK1 may also control many levels of transcription during geminivirus infection; however, the antiviral signaling pathway of SnRK1 is largely unknown.
Geminiviruses also protect some unstable host proteins such as GRIK and SAMDC1 from degradation (Shen and Hanley-Bowdoin, 2006; Zhang et al., 2011) and utilize the ubiquitin pathway for viral replication (Alcaide-Loridan and Jupin, 2012). They hijack the ubiquitin ligase complexes that are key regulators of several processes, including the cell cycle, for modulating host function. The C2 protein changes several plant hormone responses using the CUL1-based SCF ubiquitin E3 ligases (Lozano-Duran et al., 2011), and C4 activates plant cell proliferation using RKP ligase that targets cyclin kinase inhibitors for proteasomal degradation (Lai et al., 2009). In addition, the Clink protein bound to RBR and SKP1 was supposed to alter ubiquitination to affect cell cycle regulation (Aronson et al., 2000). Indeed, during geminivirus infection, there was a general increase in the transcription of genes encoding components of the ubiquitin-proteasome pathway and ubiquitin enzymes (Ascencio-Ibanez et al., 2008).
Remorins are plant-specific proteins, first reported as plasma membrane (PM) proteins in leaves of tomatoes and potatoes phosphorylated in the presence of polygalacturonide (Farmer et al., 1989). Some of them have been found in detergent-insoluble membranes called lipid rafts (Laloi et al., 2007; Lefebvre et al., 2007; Mongrand et al., 2004). Remorins, found in all land plants, are a family of proteins comprised of six different groups (Raffaele et al., 2007). Remorin genes are expressed in diverse tissues, such as embryonic, shoot apex, and vascular tissues (Bariola et al., 2004), and are induced in dehiscent tissues, source parts of the leaves, and aging organs of tobacco (Raffaele et al., 2009b). Furthermore, mRNA and protein levels of some remorins are regulated by various abiotic stressors, hormones and pathogens (Coaker et al., 2004; Li et al., 2012; Nohzadeh Malakshah et al., 2007; Raffaele et al., 2007; Widjaja et al., 2009).
Remorin proteins have a variable N-terminal, a conserved C-terminal (Remorin_C; PF03763), and a Pfam domain. The N-terminal is responsible for structural and functional divergence, whereas the C-terminal, which includes the coiled-coil motif, is important for oligomerization and localization in the PM. Recently, it was shown that the 28-amino acid C-terminal of StREM1.3, the remorin C-terminal Anchor (RemCA), was necessary and sufficient for PM targeting (Perraki et al., 2012).
Studies on plant-virus and plant-microbe interactions have reported a variety of functions for remorin. Researchers have shown that Potato StREM1.3 binds cell wall-derived galacturonides (Reymond et al., 1996) and interacts with the viral protein TGBp1 of potato virus X (Raffaele et al., 2009a); REM1.3 remorin enhances susceptibility to Phytophthora infestans (Bozkurt et al., 2014); Arabidopsis thaliana remorin 1.3 (AtREM1.3) is differentially phosphorylated after treatment with a bacterial elicitor (Benschop et al., 2007; Jarsch and Ott, 2011) and AtREM1.2 has been identified as RIN4, a negative regulator of plant immunity. (Liu et al., 2009). In addition, MtREM2.2, phosphorylated by RLK, regulates bacterial infection (Lefebvre et al., 2010), and AtREM1.3 interacts with IMPa and translocates to the nucleus (Marin et al., 2012). Although remorins have been expected to function as scaffold proteins in many important signaling pathways, their role in the signaling process is remains unknown.
Since there is little information on the remorins belonging to group 4, we studied AtREM4.1 and AtREM4.2 as the first step in understanding their biological functions in Arabidopsis. We found that they are highly regulated by osmotic stress and senescence, and they are positive regulators of geminivirus infection through interacting with SnRK1.
Materials and Methods
Plant materials and virus inoculation
All Arabidopsis used in this study were of the Columbia (Col-0) ecotype. AtREM4.1-1, AtREM4.2-1 and AtREM4.2-2 mutants were obtained from the SALK (SALK_063269C, SALK_119462 and SALK_143766) and the double mutants were constructed by crossing AtREM4.1 and AtREM4.2 mutants. For the transgenic plants of pAtREM4.1::GUS and pAtREM4.2::GUS, the 1,622 base pair (bp) upstream region of AtREM4.1 and 1,544 bp upstream region of AtREM4.2 were amplified with primers (Supplementary Table 1) by using genomic DNA. These were then cloned into the gateway pBGWFS7 vector. For overexpression lines, AtREM4.1 and AtREM4.2 were inserted into the pEarleyGate102 binary vector (Earley et al., 2006), then transgenic plants were generated using Agrobacterium tumefaciens GV3101 strains containing the constructs by a floral dip method (Zhang et al., 2006).
For virus inoculation, seeds were sowed in soil for 3 days at 4°C, and plants were grown under 100 μmole/m2/s white fluorescent lights with a 16 h light and 8 h dark cycle at 22°C. The Agrobacterium strains containing BCTV, BSCTV, and control vector pMON, were inoculated in the crown of the rosette of four-week-old plants using a needle as previously described (Lee et al., 1994).
Sequence analyses
ClustalW2 (http://www.ebi.ac.uk/Tools/msa/clustalw2/) and Boxshade (http://www.ch.embnet.org/software/BOX_form.html) programs were used to align the amino acid sequences. Conserved motifs of the protein were identified using MEME (http://meme.sdsc.edu/meme/intro.html) and ELM (http://elm.eu.org/) programs. Coiled-coil motifs were predicted using Marcoil (http://bioinf.wehi.edu.au/f-olders/mauro/Marcoil/index.html). Putative phosphorylation site and PEST motifs were predicted using NetPhos 2.0 (http://www.cbs.dtu.dk/services/NetPhos/) and epestfind (http://emboss.bioinformatics.nl/cgi-bin/emboss/epestfind).
Gene expression analysis
For reverse transcription-polymerase chain reaction (RT-PCR), 2 μg of total RNA isolated from the plants using TRIzol (Invitrogen, USA) was used as a template for cDNA, and it was performed using the gene specific primers (Supplementary Table 1). The Actin-2 gene was used as a loading control for PCR. For promoter activity analysis, the T3 transgenic plants of pAtREM4.1::GUS and pAtREM4.2::GUS were treated with MES, 100 μM ABA and 150 mM NaCl at 3 h. For histochemical analysis, plants were incubated at 37°C for 4 h in the GUS staining buffer (100 mM sodium phosphate (pH 7.0), 10 mM EDTA, 0.5 mM K3[Fe(CN)6], 0.5 mM K4[Fe(CN)6], 0.1% Triton X-100) supplemented with 0.5 mM X-Gluc. Chlorophyll was cleared as previously described (Jefferson et al., 1987).
Yeast two-hybrid (Y2H) assay
Full length cDNA, such AtREM1.4, AtREM4.1, AtREM4.2 and SnRK1.2, and truncated cDNA, such as AtREM4.1N (amino acid 1-179), AtREM4.1C (amino acid 180-296), AtREM4.2N (amino acid 1-157) and AtREM4.2C (amino acid 158-275) were amplified with the primers (Supplementary Table 1), and the amplified fragments were ligated into the pGAD GH and pGBT9 BS vectors. The constructs were concurrently transformed to the pJ69-4a yeast strain, and the double transformants in pJ69-4a strains were selected as cells grown in the absence of leucine and tryptophan. The selected double transformants were transferred and grown in the absence of adenosine, histidine, leucine, and tryptophan for interaction selection.
Bimolecular fluorescence complementation (BiFC) and localization of fluorescent conjugated proteins
AtREM1.4, AtREM4.1, AtREM4.2 and SnRK1.2 were amplified with the primers (Supplementary Table 1) and inserted into the pSPYNE173 and pSPYCE(MR) vectors (Waadt et al., 2008). The constructs were transformed to GV3101 and BiFC was performed as previously described (Voinnet et al., 2003). The GV3101 strains contained each construct at an OD600 of 0.5 and were coinfiltrated with the p19 strain at an OD600 of 0.3 into 4-week-old tobacco plants (N. benthamiana). Confocal imaging was performed 3-5 days after infiltration with the LSM700 confocal laser-scanning microscope (Carl Zeiss, Germany). To analyze the localization of AtREM4.1-CFP and AtREM4.2-CFP, we used 5-day-old seedlings of Arabidopsis overexpressing AtREM4s, and the confocal images were obtained from root.
Expression and purification of recombinant proteins
AtREM4.1, AtREM4.2, GRIK1, and truncated SnRK1.2 (amino acid 1-342) were amplified with the primers listed in Supplementary Table 1. PCR products were cloned into pGEX4T-1 and transformed into Escherichia coli strain BL21(DE3)pLysS. Cells in log-phase growth were grown in the presence of 1 mM IPTG for 20 h at 20°C. Total protein was extracted using BugBuster (Novagen, USA) and purified using GST resin (ElpisBiotech, Korea).
In vitro kinase assay
In vitro kinase assay was performed as previously described (Shen et al., 2009). Recombinant proteins (250 nM) were incubated with reaction buffer (25 mM Tris-HCl, pH7.5, 10 mM MgCl2, 1 mM EGTA, 1 mM DTT, 0.2 mM cold ATP, and 0.5 μCi/μL [γ-32P]ATP) at 30°C for 1 h, and the reaction was stopped by 5× SDS-PAGE sample buffer. An equal volume of each sample was loaded and separated in 8% SDS-PAGE gels. After protein separation, the gels were lapped and 32P-labeled proteins were visualized by autoradiography on X-ray films (Fujifilm, Japan).
Cell free degradation assay
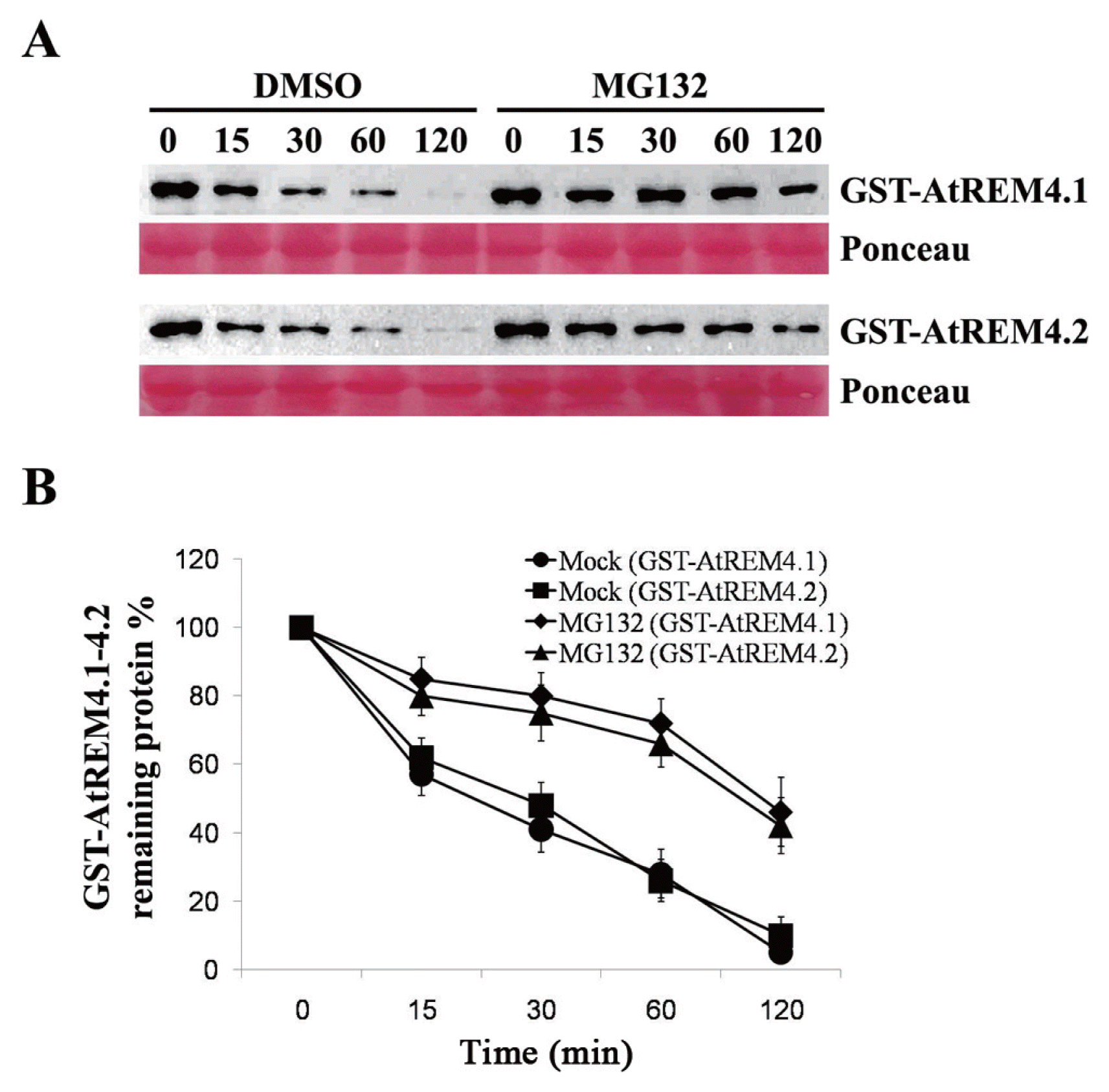
Seedling powders of Arabidopsis that were ground in liquid nitrogen were resuspended in degradation buffer containing 25 mM Tris-HCl, pH 7.5, 10 mM NaCl, 10 mM MgCl2, 5 mM DTT, 10 mM ATP and 4 mM PMSF and total proteins were extracted as previously described (Wang et al., 2009). Recombinant AtREM4 proteins were incubated in equal quantities of extracts with DMSO or 40 μM MG132 at 22°C for the indicated times, and the reaction was stop by 5× SDS-PAGE sample buffer. An equal volume of each sample was separated in 8% SDS-PAGE gel, and immunoblots were performed with GST antibody. The band quantifications of the remaining protein on immunoblots were measured using ImageJ (http://rsb.info.nih.gov/ij/index.html).
Results
Transcription of AtREM4s is highly enhanced by osmotic stress, abscisic acid and senescence
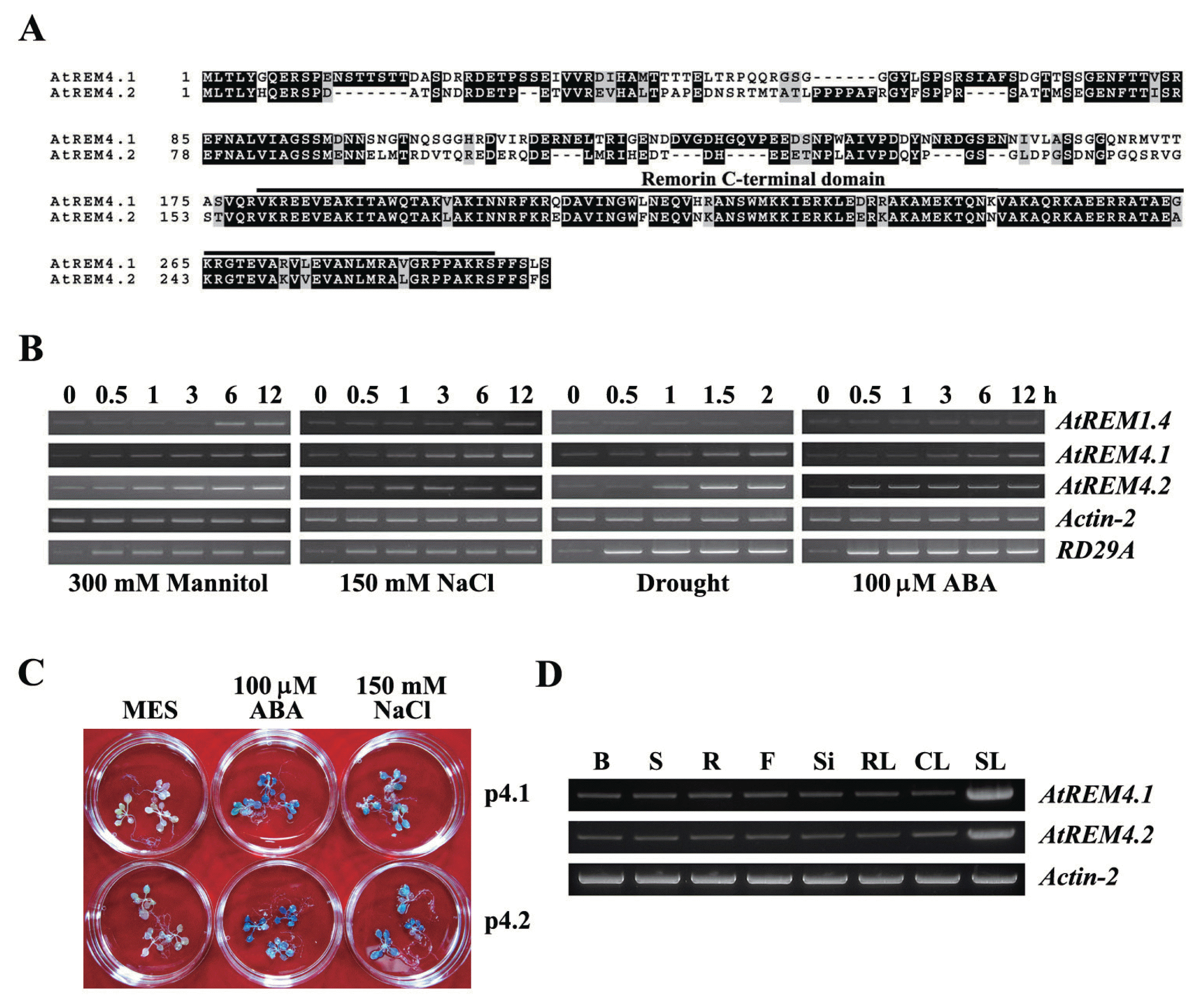
There are 16 remorin genes in A. thaliana and they are expected to be involved in various biotic and abiotic signaling pathways. However, their function is not known yet. To elucidate the role of remorin, we examined the expression levels of 16 remorin genes of Arabidopsis using the Genevestigator program (https://www.genevestigator.com/gv/) (data not shown). Interestingly, AtREM4.1 and AtREM4.2 were highly regulated by various osmotic stressors, ABA, high light, and pathogens. They have similar sequences and are likely to be recent duplicates of each other (Raffaele et al., 2007). Deduced AtREM4 proteins have proline and serine-rich N-terminal regions and each have the 89% conserved remorin C-terminal domain, a typical remorin protein structure (Fig. 1A). To validate the expression levels of AtREM4s, we performed RT-PCR using gene specific primers in various osmotic and ABA conditions. Indeed, their expressions were highly enhanced by mannitol, NaCl, drought, and ABA conditions (Fig. 1B). Moreover, their promoters were also activated strongly by NaCl and ABA (Fig. 1C). These two genes were predominantly detected in bud, stem, root, flower, silique, and leaves, and enhanced dramatically in senescence leaf (Fig. 1D).
AtREM4 proteins form homo- and hetero-interactions in the plasma membrane
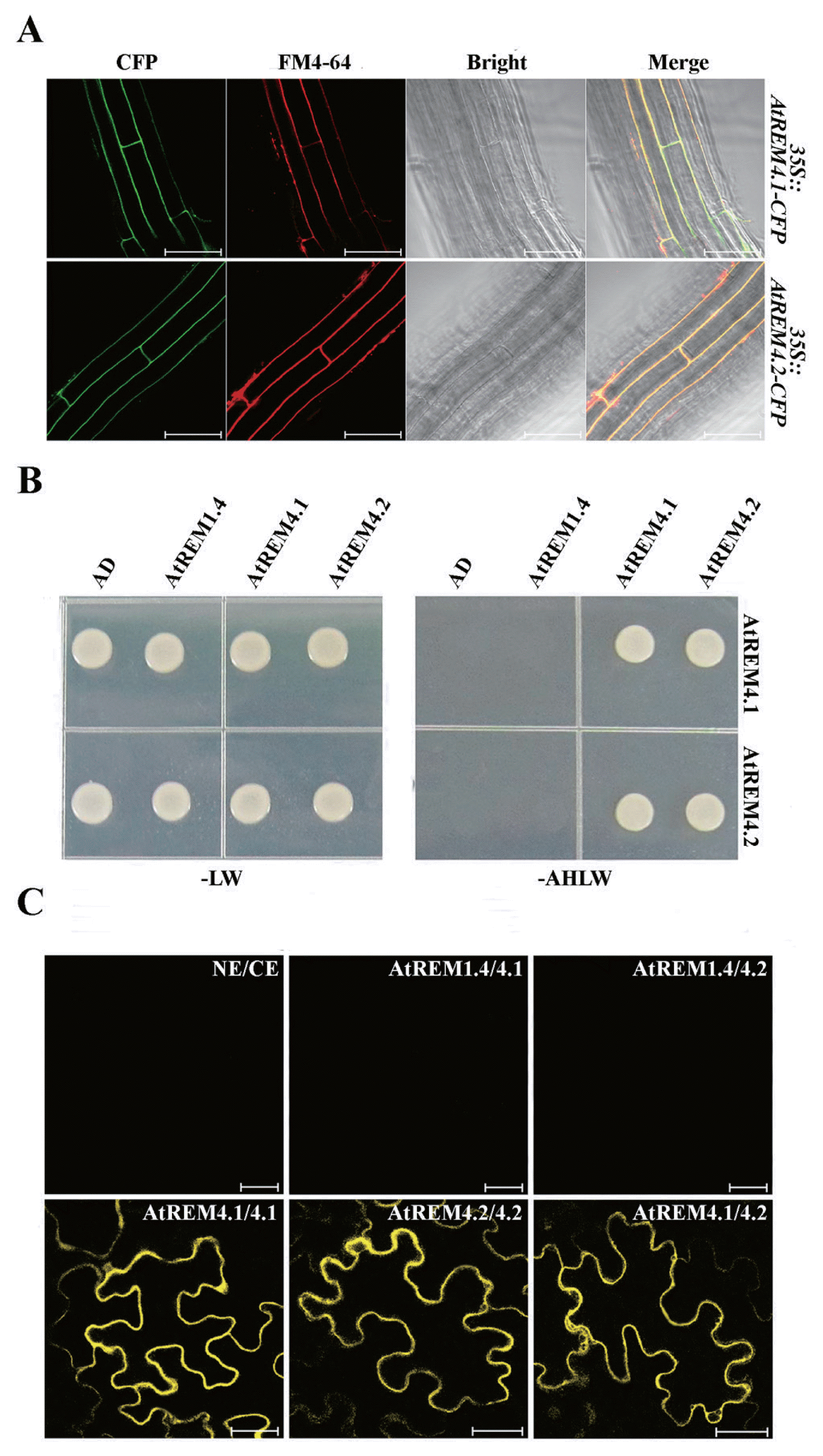
Remorin proteins are reported to localize in the PM, but localization of group 4 remorin proteins remain unknown. Therefore, in order to analyze the localization of AtREM4s in Arabidopsis, we used 35S::AtREM4-CFP transgenic plants. As a result, we could show that AtREM4s also targeted the PM (Fig. 2A). The remorin C-terminal domain, including the coiled-coil motif, was reported to mediate homo-interactions (Toth et al., 2012). Thus, we tested whether AtREM4 proteins that have similar amino acid sequences can interact with each other. In the yeast-two hybrid system, AtREM4s formed homo- and hetero-interactions, however, they did not interact with group 1 remorin protein AtREM1.4 (Fig. 2B). To confirm this result in planta, we performed BiFC using N. benthamiana leaves, where strong interactions of AtREM4s were observed in the PM of infiltrated tobacco epidermal cells (Fig. 2C).
Double mutants of AtREM4s reduce the BCTV and BSCTV susceptibility
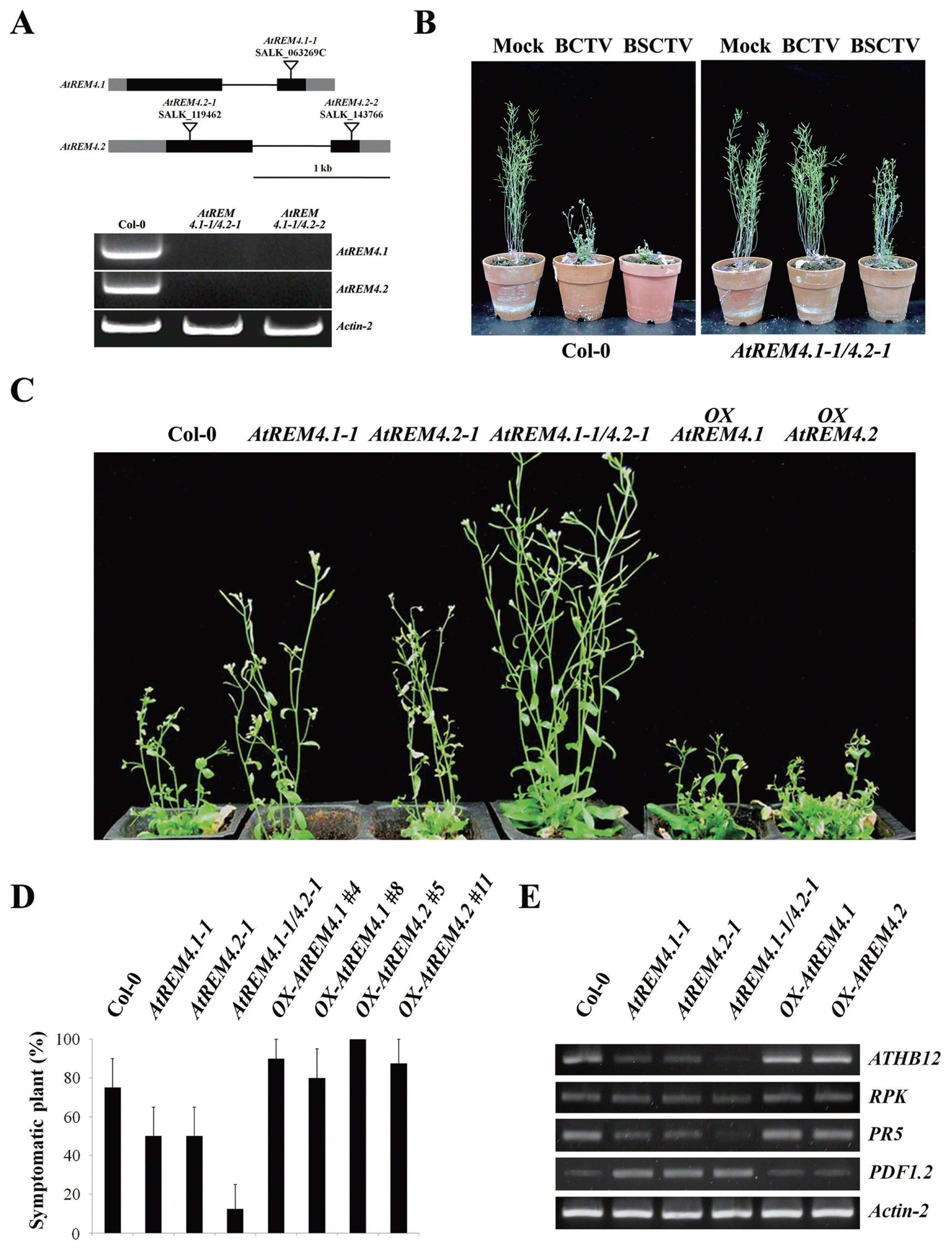
To study the function of AtREM4s, we generated double mutants (Fig. 3A) and overexpression lines (data not shown) to test their involvement in plant-microbe interaction. Interestingly, AtREM4 double mutants showed reduced BCTV and BSCTV susceptibility (Fig. 3B). To confirm this, we inoculated the Agrobacterium that contained the BSCTV genome to AtREM4 mutant and overexpression lines. As a result, single mutants showed slightly reduced susceptibility, whereas overexpression lines showed severe stunting of growth, severe curling of leaves and malformed inflorescence structures (Fig. 3C). Based on severity of infection rates, classified as previously described (Park et al., 2011), the difference was more evident (Fig. 3D). At 3 weeks after BSCTV inoculation, approximately 75% of wild-type (WT) plants developed severe symptoms. However only 10% of the plant double mutants showed severe symptoms, while more than 80% of the overexpression lines showed severe infection. Next, RT-PCR was performed to examine the infectivity at molecular level. It was reported that ATHB12 and RKP were induced by BSCTV C4 (Lai et al., 2009; Park et al., 2011). Thus, we used ATHB12 and RKP as marker genes of BSCTV susceptibility. ATHB12 and RKP were expressed highly in WT and overexpression lines, but slightly in double mutants (Fig. 3E). The gene profiling data showed that markers of SA response such as PR1 and PR5 were elevated during geminivirus infection, while transcripts for some JA markers such as PDF1.2 were reduced (Ascencio-Ibanez et al., 2008; Yang et al., 2008). Indeed, PR5 was increased more in WT and overexpression lines than mutant lines, whereas PDF1.2 higher in mutant lines than WT and overexpression lines (Fig. 3E). These results indicate that AtREM4 regulate positively the BCTV and BSCTV susceptibility.
AtREM4s interact with SnRK1.2 and AtREM4.1 is phosphorylated by it
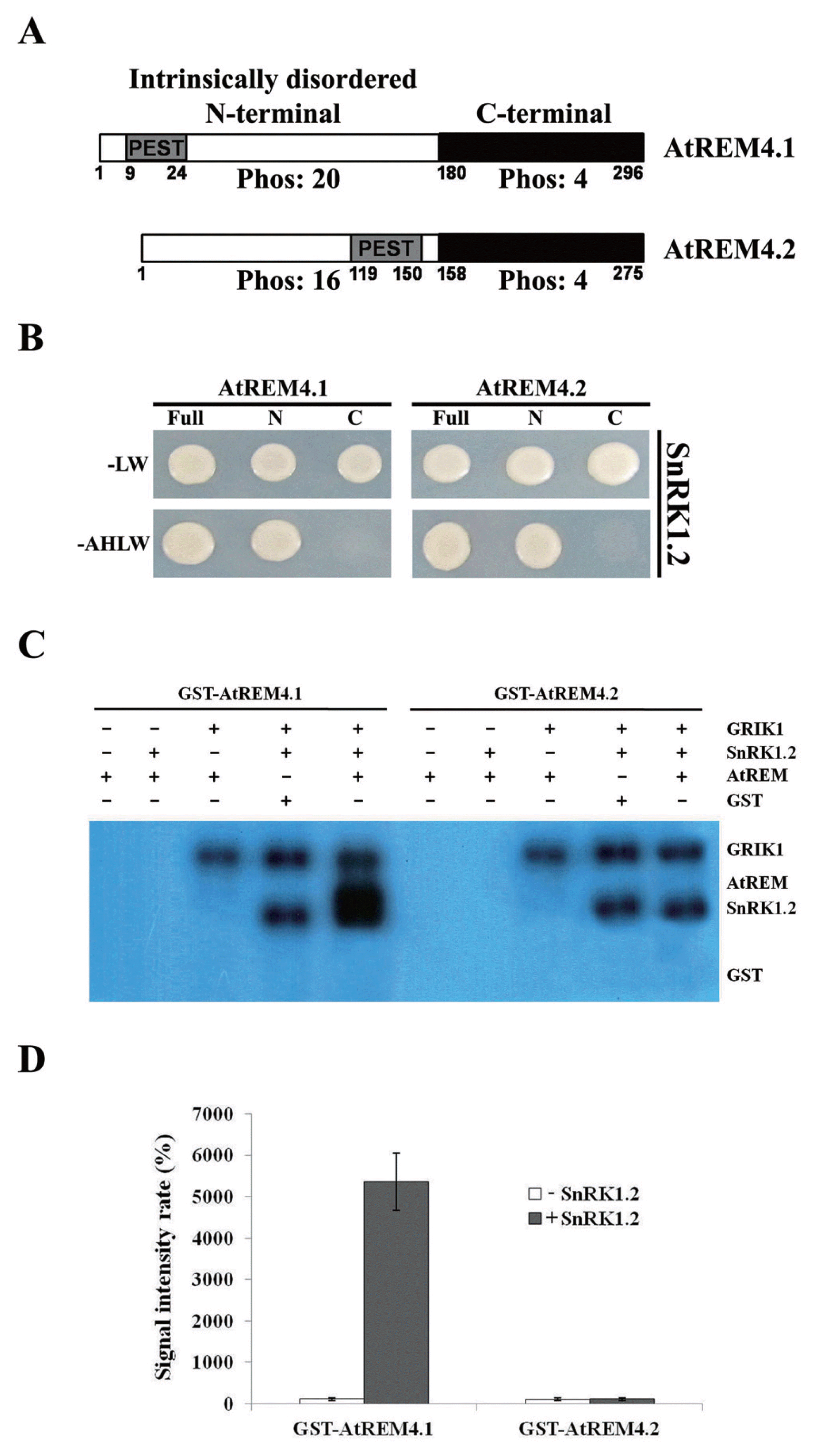
Using computer software, electrophoretic mobility, and heat stability, we found that AtREM4s have intrinsically disordered N-terminal regions (data not shown). They are flexible and modified by several different enzymes such as kinases, ubiquitin-ligases, acetyltransferases and methylases (Dyson and Wright, 2005). Indeed, a bioinformatic analysis of AtREM4s predicted putative phosphorylation sites and PEST motifs within the N-terminal regions (Fig. 4A). Using Y2H library screening, we identified AtREM4-interacting protein partners including SnRK1.2 (data not shown). AtREM4s has many serines and threonines in the N-terminal region, and the phosphorylation prediction program NetPhos2 predicted more than 20 amino acids to be phosphorylated (Fig. 4A). Thus, we examined which regions mediated this interaction using truncated versions of the gene. N-terminals of AtREM4s interacted with SnRK1.2, but C-terminals did not (Fig. 4B). To address whether AtREM4s are phosphorylated by SnRK1.2, we performed in vitro kinase assays using recombinant proteins with GRIK1, an upregulator of SnRK1 (Hao et al., 2003; Shen et al., 2011). Various combinations of recombinant AtREM4s, SnRK1.2 and GRIK1 proteins were incubated in the presence of [γ-32P]ATP, and phosphorylation was visualized by autoradiography. The signal band was detected strongly in the GST-AtREM4.1 loading line, but not in the GST-AtREM4.2 line (Fig. 4C). Quantitative analysis of the signal intensity rate also showed that only GST-AtREM4.1 shows an intense signal (Fig. 4D).
AtREM4s are degraded by the 26S proteasome pathway
Many unstable proteins that are degraded by the 26S proteasome have a PEST motif (Rechsteiner and Rogers, 1996; Rogers et al., 1986). Thus, we tested whether AtREM4 proteins are degraded in a proteasome-dependent manner. In the mock, GST-tagged AtREM4s were degraded rapidly, whereas the proteasome inhibitor MG132 apparently delayed the degradation such that with MG132 (Fig. 5A), GST-tagged AtREM4s remained 42-46% after 2 h, however, in mock, it remained only 5-10% (Fig. 5B). These results indicated that AtREM4s are degraded by the 26S proteasome.
Discussion
In this study, we present for the first time, the molecular characteristics of AtREM4s and their putative roles during the geminivirus infection process.
AtREM4s are typical remorins and their expression is highly stimulated by osmotic stress and senescence
To date, most of the studies on remorins have focused on group 1 and 2. Thus, in this study, as the first step in elucidating the biological function of remorin group 4, we studied AtREM4.1 and AtREM4.2. They showed typical characteristics of the remorin molecules, which consist of variable N-terminal and conserved C-terminal regions such as the remorin group 1 proteins (Fig. 1A). Remorins have been known to respond to various stress or developmental conditions. Transcription rates and protein levels of some group 1 remorins were regulated by salt, drought and ABA (Nohzadeh Malakshah et al., 2007; Raffaele et al., 2007), and the transcription of NtREM1.2 increased with organ aging (Raffaele et al., 2009b). AtREM4s were also regulated similarly by abiotic and biotic stressors. However, of the 16 AtREMs, the expression of AtREM4s were induced to the greatest extent by various osmotic stressors and ABA (Fig. 1B). They were also expressed strongly in senescing leaves (Fig. 1D). Moreover, AtREM4s were localized in PM (Fig. 2A) and formed homo-oligomers (Fig. 2B) such as StREM1.3, AtREM1.3 and MtREM2.2. Interestingly, AtREM4s also formed hetero-oligomers, while did not with AtREM1.4 (Fig 2B), suggesting that they could share interacting components each other, and play redundant roles in the same signaling pathway.
AtREM4s enhance susceptibility to geminivirus by mediating SnRK1 and RBR interaction
In this study, we show that AtREM4s enhance susceptibility to geminivirus: overexpressors showed severe infection, while double mutants showed resistance (Fig. 3B). Interestingly, AtREM4s have putative motifs such as the FHA, BRCT and WW domains found in regulating complexes of cell division and DNA damage repair proteins. Geminivirus infection leads to cell cycle reprograming of host plants and activation of G1/S transitioning for geminivirus DNA replication. RBR is a key negative regulator of that transition in plant cells (Gutzat et al., 2012). It binds and inactivates E2F transcription factors required for the expression of genes such as the host replication initiator proteins (Rep). Indeed, interaction of Rep with RBR led to the development of geminivirus symptoms (Kong et al., 2000). Moreover, we show that AtREM4s interacted with SnRK1 (Fig. 4B), by which AtREM4.1 was phosphorylated in vitro (Fig. 4C). GRIK, identified as Rep interacting protein, (Kong and Hanley-Bowdoin, 2002) phosphorylated SnRK1 (Shen et al., 2009). SnRK1, a key regulator of plant stress and metabolism, is involved in plant antiviral defense. Overexpression of SnRK1 resulted in reduction of geminivirus infection, while silencing of it resulted in the accumulation of more viral DNA than wild-type plants (Hao et al., 2003; Shen et al., 2011). These results suggest that AtREM4s may be one of the intermediate regulators between SnRK1 and RBR during geminivirus infection. Alternatively, they might be involved in a SnRK1-mediated defense-signaling pathway. To prove this hypothesis, we are in the process of elucidating the functional roles of AtREM4 and AtREM4-interacting partners using genetic and cell-biological approaches.
In addition, AtREM4s interacted with some ubiquitin-proteasome components (data not shown) and AtREM4.1 was degraded by the 26S proteasome (Fig. 5A). Previously, there were some reports that remorin protein levels were increased, while the expression was not changed or even decreased (Nohzadeh Malakshah et al., 2007; Widjaja et al., 2009). When we searched the PEST motifs, known as a signal for protein degradation, among 16 remorin proteins in Arabidopsis, they were predicted in N-terminals of 14 remorins except AtREM3.1 and AtREM3.2 (data not shown). These results suggest that decreased protein levels of the remorins mediated by the 26S proteasome, such as degradation of AtREM4 or AtREM4-interacting protein during the geminivirus infection process, might also be an important mechanism in regulating remorin-mediated signaling processes.
Supplementary Table
Arabidopsis thaliana Remorins Interact with SnRK1 and Play a Role in Susceptibility to Beet Curly Top Virus and Beet Severe Curly Top Virus.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement
Supplement Print
Print






