

The filamentous fungus Curvularia lunata (Wakker) Boedijn (telemorph: Cochliobolus lunatus R. R. Nelson & F. A. Haasis) is a ubiquitous plant pathogen, which is causal agent of Curvularia leaf spot (CLS) (Macri & Lenna, 1974). Infection of maize leaves with C. lunata leads to huge yield loss in severe epidemic years in northern China. For instance, it occurred over 192,000 hm2 and led to 8 million kg yield loss in Liaoning province in 1996 (Dai et al., 1995; Dai et al., 1998). Because of nationwide extensive application of resistance varieties containing tropic and sub-tropic germplasms, the incidence of disease infection and its severity were declined massively and less damage was observed in field. However, in recent years, the disease has bounced back again and caused serious damages in some maize growing areas such as Liaoning, Henan and Anhui province etc (Gao et al., 2014b). More importantly, methyl 5-(hydroxymethyl) furan-2-carboxylate (M5HF2C) produced by C. lunata in infected grains pose a serious threat to human and animal health (Gao et al., 2014a). Despite the high economic impact of CLS, efficient strategies for the management of CLS have not been fully developed, which could result partially from our limited knowledge on C. lunata pathogenomics.
In the last two decades, significant progress has been made towards a better understanding of the factors involved in the pathogenesis of C. lunata. A multiple virulence factors have been demonstrated to be involved in pathogen infection to maize, such as cellulose (Feng et al., 2002), non-host specific toxin (methyl 5-(hydroxymethyl)-furan-2-carboxylate) (Liu et al., 2009), melanin (Xu et al., 2007). It is worth mentioning that some of virulence related genes have been successfully cloned in previous work such as Brn1 being required for DHN melanin synthesis (Liu et al., 2011), Clt-1 regulating non-host specific toxin production (Gao et al., 2015), two mitogen-activated protein kinases (MAPK) encoding genes (Clk1 and Clm1) (Gao et al., 2012; Wang and Chen, 2011). More recently, it has been reported that the Brn1 gene is not only responsible for melanin synthesis but also contributes, to some extent, to toxin production. Whereas, the related pathogenicity genes in melanin and toxin production have not been extensively studied.
A proteomic approach allows the simultaneous characterization of a large number of relevant proteins expressed by a genome (Conrad et al., 2008). Using the proteomic approach, researchers have successfully identified a Clt-1-regulated protein scytalone dehydratase (SCD, accession: KJ507954) associated with the production of toxin and pathogenicity in C. lunata (Gao et al., 2015). These studies are crucial in understanding the regulating network underlying pathogen virulence. However, the information presented in these studies can not directly reveal the proteins action mechanism of disease’s pathogenicity. Information on pathogenic factor interrelation and their interactions on each other is required for developing strategies to prevent fungal infection. Protein interaction networks are of fundamental importance to almost all biological processes, a powerful and high-throughput protein-protein Y2H screen system becomes more and more crucial to identify or mine the partners of proteins in the regulatory complexes (Miller and Stagljar, 2004).
To screen the pathogenesis-related interaction protein, an efficient and high-quality full-length cDNA library is necessary for studying the protein function and finding the protein associated with the pathopoiesia mechanism against host leaf. This study described a method for constructing a full-length cDNA library of C. lunata using the switching mechanism at the 5′-end of RNA transcript (SMART) technology. Moreover, melanin synthesis protein 1,3,8-hydroxynaphthalene reductase (Brn1 gene, GeneBank accession number: DQ358052) was used as a “bait” to test the efficiency of the Y2H library. This study not only demonstrated the critical techniques to construct a high-quality Y2H library but also provided informative cues to unravel interacting proteins with Brn1 involved in multiple secondary metabolic pathways in C. lunata.
Materials and Methods
Fungal strains used for the Y2H library
C. lunata CX-3 strain was selected for constructing cDNA library since it is highly virulent to maize and caused typical lesion on the maize leaves. The monoconidial culture of C. lunata was grown on potato dextrose agar (PDA) medium at 28°C (Gao et al., 2014a). The strain is stored in our laboratory and used for the study of pathogenicity mechanism of the pathogen for 10 years. The strain is maintained on PDA medium at 4°C or silicone beads at −20°C.
cDNA library primer
Universal primers were designed to amplify the inserted fragments of the cDNA library according to vector pGADT7-Rec sequence, and synthesized by Shanghai Sangon Company (Shanghai, China) as follows: MATCHMAKER 5′ AD LD-Insert Screening Amplimer: 5′-CTATTCGATGATGAAGATACCCCACCAAACCC-3′; MATCHMAKER 3′ AD LD-Insert Screening Amplimer: 5′-GTGAACTTGCGGGGTTTTTCAGTATCTACGATT-3′.
Total RNA isolation
To extract fungal RNA, the strains were grown in 100 ml of PD medium in 250-ml Erlenmeyer flasks with continuous shaking at 160 rpm for 72 h at 28°C. The mycelia produced from these cultures were harvested by filtration using three layers of sterile cheese cloth, washed with sterilized double-distilled (dd)H2O, and ground in liquid nitrogen. Total RNA was extracted using Trizol Reagent (Invitrogen, Carlsbad, CA) from the powdered mycelia according to the manufacturer’s instructions, and then treated with DNaseI (Takara, Japan) to remove eliminate contaminated genomic DNA. The integrity of total RNA was analyzed by 1% agarose gel electrophoresis, followed by ethidium bromide (EtBr) staining. The concentration and purity of total RNA were determined by a spectrophotometer (Eppendorf AG, Hamburg, Germany) at 260 and 280 nm, respectively.
cDNA synthesis
The single-stranded cDNA (sscDNA) were synthesized using a SMARTTM cDNA Library Construction Kit (Clontech USA, Mountain View, CA) according to the manufacturer’s instructions. sscDNA were synthesized by 1 μg of total RNA using SMART III oligo (5′-AAGCAGTGGTATCAACGCAGAGTGGCCATTATGGCCGGG-3′), CDS III primer (5′-ATTCTAGAGGCCGAGGCGGCCGACATG-d(T)30VN-3′), and Moloney Murine Leukemia Virus (MMLV) Reverse Transcriptase. Double-stranded cDNA (dscDNA) were acquired by 20 cycles of long-distance PCR using Advantage 2 Polymerase Mix (Clontech USA, Mountain View, CA) with 2 μl of sscDNA product. The reaction parameters were as follows: pre-thermal denaturation at 95°C for 30 s; followed by 20 cycles of 95°C for 10 (extension/enhance 5 s after each cycle), and 68°C for 6 min. To identify the quality of dscDNA, 5 μl of the PCR products was analyzed on 1% agarose gel electrophoresis, followed by EtBr staining. To get rid of low-molecular-weight dscDNA fragments, small DNA contaminants, and unincorporated nucleotides from dscDNA, the PCR products were purified by a CHROMA SPINTM TE-400 column (Clontech USA, Mountain View, CA). The purified dscDNA were quantified by electrophoresis on 1% agarose/EtBr gel.
Construction and evaluation of the cDNA library
The purified dscDNA, together with linearized pGADT7-Rec AD cloning vector (Clontech USA, Mountain View, CA), were co-transformed into yeast competent cell AH109, where yeast repair enzymes restore the linearized plasmid to its circular form by recombining homologous sequences at the end of the dscDNA and pGADT7-Rec. The well-distributed yeast cells were then cultured on 100 dropout media (SD/-Leu) 150 mm plates, and incubated at 30°C until clones appeared (3-6 d). All clones were pooled with freezing medium (YPDA medium with 25% glycerol) and stored at −80°C. Transformed yeast cells (1 ml) were diluted with YPDA medium, and 100 μl was spread in dilutions of 10−1, 10−2, 10−3, 10−4, and 10−5dilutions on 150 mm SD/-Leu plates, respectively. After culturing at 30°C for 3-6 d, the positive transformants were harvested to form an Y2H library. The transformation efficiency and library titer were calculated by counting the clones (cfu) as follows: transformation efficiency = Colonies on SD/-Leu × dilution factor ÷ volume (ml) plated × 6 ml per 3 μg pGADT7-Rec; library titer (cfu/ml) = Colonies on SD/-Leu × dilution factor ÷ volume (ml) plated. The colonies were randomly selected to identify the size of the inserting cDNA fragment and calculate the reorganization ratio using MATCHMAKER AD LD-Insert primers based on the following PCR conditions: 94°C for 3 min; 30 cycles of 94°C for 30 s, 55°C for 30 s, 72°C for 1 min; and 72°C for 10 min. The PCR product (5 μl) was load on a 1% agarose/EtBr gel. Recombination rate = reaction number with cDNA fragments/total reaction number × 100%.
C. lunata Y2H library application for the Brn1 protein interaction
In this study, Brn1 (melanin synthesis protein 1,3,8-hydroxynaphthalene (THN) reductase) was selected to detect the sufficiency of the resultant Y2H library. The coding sequence of Brn1 encoding 267-amino acid were amplified from the cDNA of C. lunata. The cDNA fragment of Brn1 was inserted into the EcoRI-BamHI sites of the yeast GAL4 binding domain vector pGBKT7 (Clontech, Mountain View, CA, USA) to generate BD-bait plasmid (pGBKT7-Brn1). Subsequently, the fusion construct was transformed into the yeast strain Y187 and the positive colonies were selected on SD/-Trp medium. Furthermore, the toxicity and autotranscriptional activation of the fusion construct were tested on SD/-Trp/+Kan (20 ug/ml) liquid medium and SD/-Ade/-Trp/+X-α-GAL plate, respectively. The Y2H AD fusion library was screened using the pGBKT7-Brn1 vector in yeast-mating way. The C. lunata library strain (AH109) with Brn1-bait-expressing reporter strain (Y187) were co-cultured, without time-consuming and labor-intensive plasmid extraction as well as co-transformation steps required in traditional Y2H screens. A clone from the bait transformant was mated with a clone from the prey transformant, and grown at 30°C overnight in 1 mL of yeast extract peptone dextrose broth. The mated clones were selected on SD medium fortified with both tryptophan and leucine to ensure successful mating. Finally, the interacting partners were screened on SD/-Leu/-Trp/-His/-Ade/+X-α-Gal.
Direct-Y2H assay
To test the auto-activation or interaction, the prey cDNA clones were co-transformed with either empty pGBKT7 vector or bait construct into AH109 strain. The co-transformants were first selected on SD/-Leu/-Trp plates and later tested for the reporter gene expression on SD/-Leu/-Trp/-His/-Ade/+X-α-Gal plates.
Results
Total RNA analysis
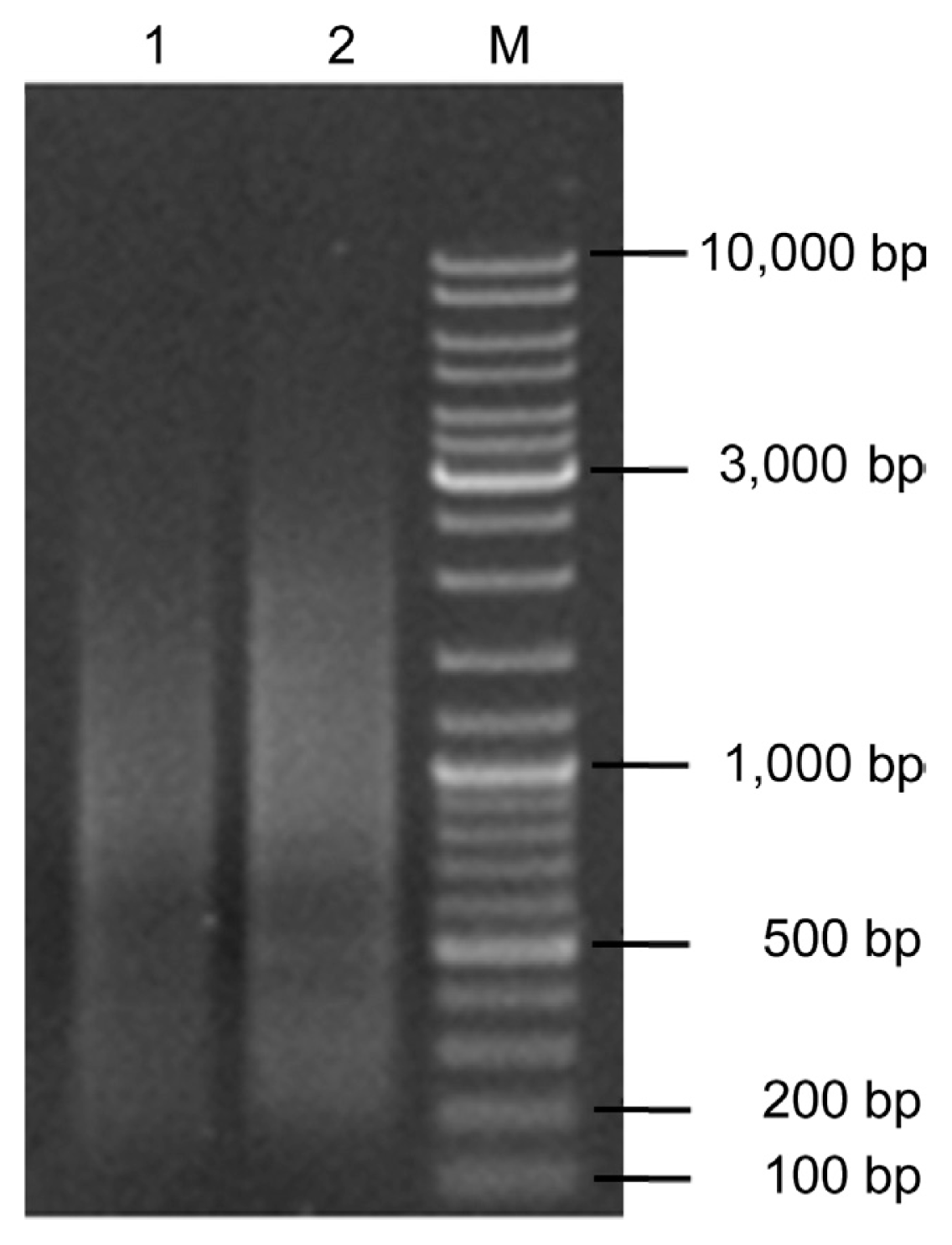
The integrity of the total RNA, extracted with the Trzol reagent from C. lunata, was identified by 1% agarose gel electrophoresis. It showed three bands corresponding to ribosomal 28S, 18S and 5S RNA, and there was no visible degradation (Fig. 1). The OD260/OD280 ratio of total RNA was 1.94 with a concentration of 728.6 ng/μL, which indicated that the total RNA had high purity and quality to construct the cDNA library.
dscDNA synthesis
dscDNA (5 μl) was analyzed by electrophoresis on 1% agarose gel. Results showed that the bands of dscDNA fragments ranged from 0.1 kb to 4.0 kb in size (Fig. 2). After purification using a Chroma Spin TE-400 column, the fragments smaller than 200 bp were eliminated, which could effectively avoid the very small inserts and non-recombinant clones in the library (Fig. 2).
Construction and evaluation of the Y2H cDNA library
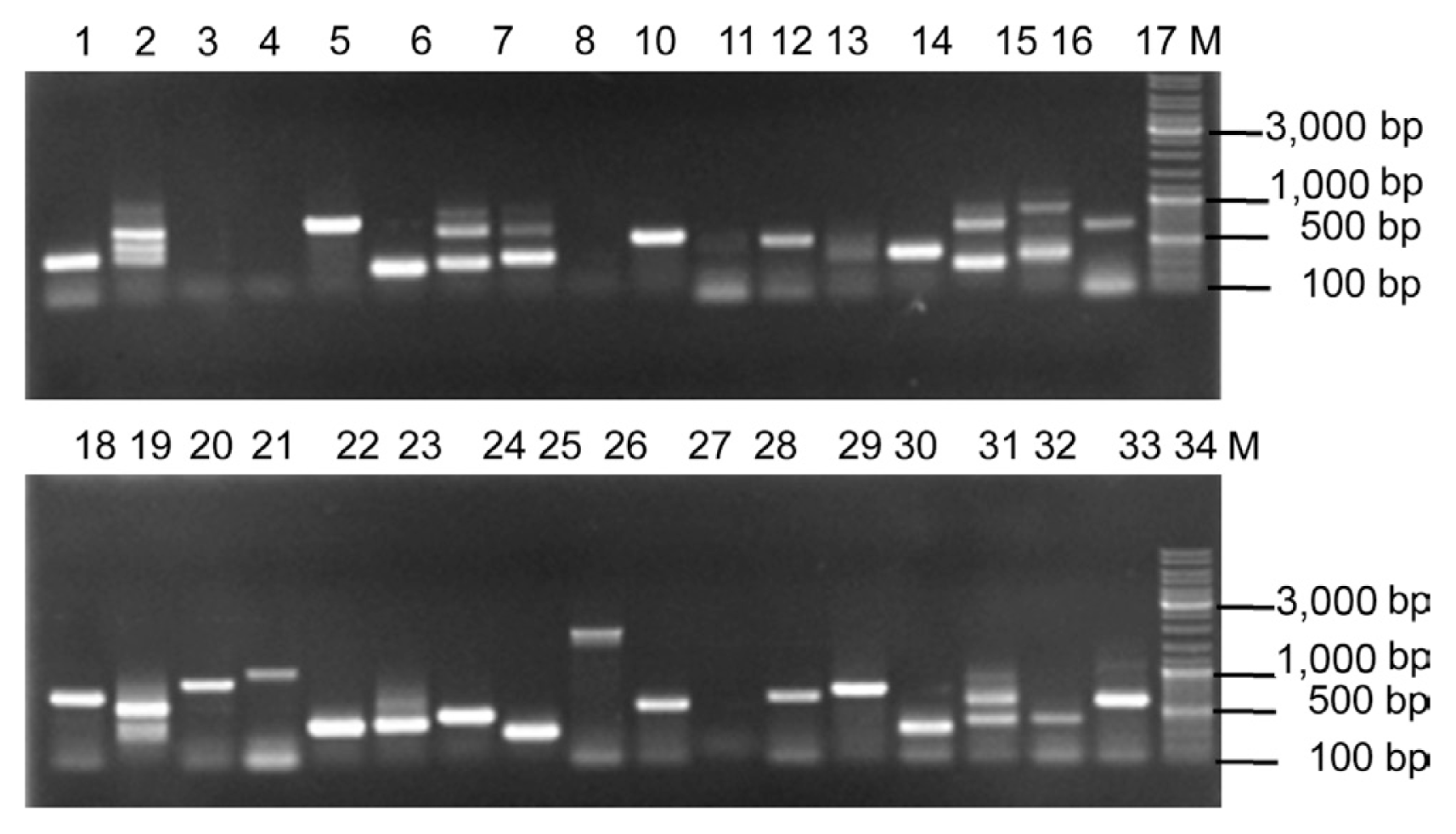
An Y2H cDNA library was constructed. Based on the clone number on SD/-Leu plates, the transformation efficiency was 6.39×105/3 μg PGADT7-Rec and the AH109 library titer was 2.5×108 cfu/ml, and the quantity of the cDNA library was 2.46×105. In order to identify the length of the inserts and the recombination rate of the cDNA library, 34 positive clones were randomly selected and identified by PCR using the MATCHMAKER AD LD-Insert primers. Results showed that most of the insert ranging from 0.4 to 3.0 kb (Fig. 4). Most of the lanes appeared as a single band. Randomly picked clones show that the recombination rate was 88.24%. The above information also indicated that the small fragment cDNA was removed from the library, and the cDNA library could be used for further research. The Y2H cDNA library was harvested with freezing medium and stored at −80°C for further use in Matchmaker TM gold Y2H system.
Quality test of Y2H screens
A total of 2.46×105 clones (C. lunata cDNA Y2H library) were screened using Brn1 ORF as “bait” (Fig. 4). Among them, 3 independent clones that fulfilled the criteria of interaction with Brn1 ORF were obtained and sequenced. The sequence similarity analysis showed they were VelB, general amino acid permease Agp2 and GTPase activating protein Sec23a. To confirm interactions between Brn1 and its partners of interest, we co-transformed the corresponding DNA-binding domain (BD) and transcription-activating domain (AD) plasmids into yeast and re-identified on QDO/X-α-Gal. The results confirmed these proteins had genuine interactions in living cells of yeast.
Discussion
The Y2H technology is by far the most powerful and widely used method to detect the protein-protein interactions in high-throughput screenings of proteomics. (Stynen et al., 2012). A high-quality expression cDNA library can provide molecular resources for analyzing the genes involved in biology and studying their protein functions and interactions facilitate the constructing of the protein network stemming from a known protein. cDNA libraries have been constructed for many plant species (Lee et al., 2012; Ma et al., 2009), animals (Han et al., 2008; Ma et al., 2011) and microorganisms (Wang and Chen, 2011; Zhao et al., 2009) and genes related to biological processes have been determined. Also, genes related to the biology of fungal pathogens, such as pathogen invasion, development, survival, pathogenicity and virulence can be discovered with a cDNA library. Few reports have been published concerning the construction of a cDNA library from C. lunata. To study the pathogenesis-related protein interaction, a full-length cDNA library from C. lunata mycelia and applied it to Y2H technology.
Conventional cDNA library construction methods suffer from several major shortcomings (Gubler and Hoffman, 1983). For example, the majority of cDNA clones lost whole or small fragments, especially mRNAs longer than 2 kb. Directional cloning suffers from methylation and is often incomplete in protecting the internal restriction sites and inefficient for cloning (McClelland et al., 1994). Given the low efficiency of enzymatic enrichment steps, conventional cDNA library construction methods are also unsuitable in cases with limited starting material. As a novel and useful method for constructing a full-length cDNA library, SMART technique increases the lengths of cDNA clone and high-quality cDNA library by simultaneously employing two intrinsic properties of MMLV reverse transcriptase (template switching and reverse transcription) (Kulpa et al., 1997; Zhu et al., 2001). This method also anchors both ends of sscDNA by adding a distinct SmaI site to each end during reverse transcription. This novel integration of template switching with SmaI-based anchoring and directional cloning ensures the construction of cDNA libraries with high yields of representatives. To perform a comprehensive screen for positive interactors, it is critical that the library have as high a complexity as possible to maximize the chances of finding relevant prey proteins. Our cDNA library contained >2.5×108 cfu/mL with an average insert size of 0.4-3.0 kb, thereby ensuring a complete representation of rare sequences.
Fungal melanins are black or dark brown pigments formed by the oxidative polymerization of phenolic compounds (Bell and Wheeler, 1986). They are not considered essential for growth and development of fungal cells, but they enhance the survival and competitive abilities of fungi in certain environments (Butler & Day, 1998). Melanin appears to contribute to virulence by reinforcing the mechanical strength of the infection into host plant epidermis in many plant diseases (Kawamura et al., 1999; Brakhage et al., 1999; Liu et al., 2011). In the maize pathogen C. lunata, the melanized layer of its appressoria and infection pegs, is essential for maintenance of the turgor pressure that enables effective leaf penetration. Melanin in conidia in nature primarily via a polyketide synthetase pathway from 1,3,8-trihydroxynaphthalene (THN) reductase gene Brn1, associated with the melanin pathway of C. lunata, was identified and characterized in our previous study (Liu et al., 2011). The Brn1 gene may also be associated with toxin production and conidiation. However, there is no detailed knowledge to reveal the genetically regulation mechanism of those virulence factors in the C. lunata so far. For this reason, we searched for more pathogenic factors, from which we could obtain more regulators that mediate melanin and toxin production, to obtain a better understanding of the regulation network that controls secondary metabolite production and conidiation.
In this study, Brn1 ORF was selected as “bait” to screen the C. lunata cDNA library. Finally, 3 proteins with positive interactions with Brn1 ORF were confirmed by sequence alignment and database retrieval. They are VelB, general amino acid permease AGP2, and GTPase activating protein Sec23a. Among these interacted proteins, VelB is our most interested protein. It carries two velvet domains located in the N- and C-terminal half, respectively. Velvet domain which represents a protein-protein interaction domain is found in filamentous ascomycetes and basidiomycetes (Bayram & Braus, 2008). VelB is involved in the regulation of diverse cellular processes by interacting with different proteins. VelB had been described as a light-dependent regulator of fungal development and secondary metabolism of A. nidulans, which interacts with VeA. VelB can also form homodimers and is part of another heterodimer, VosA-VelB. VosA-VelB represses asexual development and is required for the viability of spores by activating trehalose biogenesis. In a submerged culture during vegetative growth, VosA-VelB can negatively regulate asexual conidiation and support trehalose biosynthesis (Bayram et al., 2012).
Brn1 has long been known to be closely related to the biosynthesis of melanin. However, our previous studies showed that Brn1-silenced transformants have significantly reduced toxin production from C. lunata. Our experiment of transcriptome analysis with consecutively induced C. lunata strains on resistant varieties also demonstrated that the strains with improved virulence could simultaneously up-regulate some gene expression like melanin synthesis genes, e.g. Brn1 and SCD genes, toxin production related genes, e.g. aflatoxin biosynthesis protein ord2 gene and HC-toxin synthetase gene (data not shown). Thus it is clear that Brn1 gene in C. lunata is closely associated with toxin biosynthesis (Liu et al., 2011). VelB is known with certain relation to fungal toxin production like sterigmatocystin (ST) in Aspergillus parasiticus, cyclopiazonic acid, aflatoxin in Aspergillus flavus, ST and β-l-catam in A. nidulans, and deoxynivalenol (DON) mycotoxin in Fusarium graminearum (Jiang et al., 2012). Therefore a question should be answered whether there is a co-regulation between Brn1 and VelB. If the co-expression of Brn1 and VelB genes genuinely exists, it would be rational to consider melanin and toxin as interrelated components of a virulence entirety. The new finding of VelB associated with Brn1 would be helpful for us to globally understand the pathogen virulence mechanism.
Currently, the pathogenesis of C. lunata is not very clear. In particular, the molecular interactions of pathogenesis-related proteins are not yet fully understood. In the current study, the Y2H system was first successful constructed and adopted to screen pathogenesis-related proteins that interact with Brn1. The exact role of C. lunata VelB in pathogenicity still needs to be determined by gene function analysis. Therefore, this study could serve as the starting point for a deeper analysis on the pathogenicity in C. lunata. The elucidation of this protein interaction is of much importance both in understanding the regulatory network of melanin and toxin synthesis, and as potential targets for further research on the efficient strategies for the management of CLS.


 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






