

Bacterial ring rot, which is caused by the gram-positive bacterium Clavibacter michiganensis subsp. sepedonicus (Cms), occurs throughout potato-growing regions worldwide (Kado, 2010). This pathogen is mainly transmitted through noncertified potato seed pieces that were handled without care during the preparation of seed cutting (Manzer and Genereux, 1981). Improper sanitation methods that are used during cool temperature storage and in planting machinery can also spread the pathogen to new seed pieces (Bonde, 1942). Thus, Cms cells are easily transferred to new sites, resulting in the classification of this pathogen as highly contagious. Because of the highly contagious nature of the ring rot pathogen, many countries have implemented a quarantine program with a zero-tolerance policy (Gutbrod, 1987). In order to agree with this policy, seed and plant bioassays have firstly required the global adoption and acceptance of rapid and reliable tests by independent laboratories or institutions, including tests for new strains and hosts. Nevertheless, some of these assays cannot detect the pathogen and have some limitations.
In general, the diagnosis of Cms is based on the isolation of the pathogen, followed by Gram staining combined with a bioassay and morphological and biochemical characteristics and/or semi-selective medium and pathogenicity tests (Bishop and Slack, 1987; Cruz et al., 1992). Such tests require one to several weeks before final confirmation is obtained. Currently, serological and molecular assays based on immunofluorescent-antibody staining (IFA), enzyme-linked immunosorbent assay (ELISA), and PCR are widely used to detect Cms strains; however, because these assays also detect other closely related bacterial strains, there have been critical defects in the diagnosis and identification of Cms isolates (De Boer and Wieczorek, 1984; De Boer et al., 1988; Hu et al., 1995; Lee et al., 1997; Li and De Boer, 1995; Li et al., 2006; Mackay et al., 2002; Mills et al., 1997; Pastrik, 2000; Rademaker and Janse, 1994; Schaad et al., 1999; Schneider et al., 1993). Previous studies have developed a real-time PCR method using the cellulase A (CelA) gene from a small cryptic plasmid (pCS1), but plasmid-based genetic sequences are still quite variable (Gudmestad et al., 2009).
Recently, a number of bioinformatics applications have been proposed or innovatively used to identify discriminatory bacterial species- and subspecies-specific markers for bacterial detection and identification (Albuquerque et al., 2009; Chen et al., 2010; Cho et al., 2011; Lang et al., 2010).
In this study, Cms89F/R primer was identified using a comparative analysis of the subspecies group of genome sequences of C. michiganensis. Additionally, a SYBR Green qPCR assay was developed to detect Cms based on Cms89F/R. The efficiency, specificity and sensitivity of putative phage-related proteins were evaluated, and the efficacy of the analytical method to detect Cms in both symptom and symptomless tubers was demonstrated.
Materials and Methods
Bacterial strains and culture conditions
The bacterial strains that were used in this study were obtained from the Belgian Coordinated Collections of Microorganisms (BCCM), the Korean Agricultural Culture Collection (KACC), and the National Collection of Plant Pathogenic Bacteria (NCPPB) in the United Kingdom. Bacterial strains are listed in Table 1. The culture media and incubation conditions were taken from the Handbook of Microbiological Media (Atlas, 2004). The Clavibacter species were grown on yeast extract glucose mineral salts agar (YGM-Agar) at 23°C.
DNA extraction
Clavibacter strains were cultured on YGM medium and harvested with a scraper for total genomic DNA extraction. The total genomic DNA from the other microorganisms was prepared using a DNeasy Tissue kit (QIAGEN, Hilden, Germany) according to the manufacturer’s instructions. A Nanodrop® ND-1000 Spectrophotometer (NanoDrop Technologies, Wilmington, DE, U.S.A.) was used to measure the genomic DNA concentration. The total genomic DNA of the bacterial strains was diluted in TE buffer (pH 8.0) to 25 ng/μl.
Comparative genomic analysis and primer design
The genomes of C. michiganensis subspecies were compared using bioinformatics tools. A search algorithm was used to identify orthologues among Cms, C. m. subsp. michiganensis and C. m. subsp. nebraskensis. The data on the genomic sequences and annotations were obtained from the NCBI GenBank database (ftp://ftp.ncbi.nlm.nih.gov/genomes/bacteria/). The candidate target genes were selected using a method that was modified from Chen (Chen et al., 2010) and Lang (Lang et al., 2010). The NCBI programs BLASTN, BLASTX and Conserved Domains were used to conduct similarity and domain searches of the candidate target genes. The candidate target genes of Cms ATCC 33113 were screened by comparative genomics. In particular, the putative phage-related protein (GenBank accession No. CAQ00928, gi|169155030:861988-863124) was selected among the candidate genes of the Cms ATCC 33113 genome. The oligonucleotide sequences of the primer sets are presented in Table 2. The primers and probes for the qPCR were synthesised by Bioneer Corporation (Daejeon, Korea), respectively.
Conventional and real-time PCR conditions
The thermocycling conditions that were used for the conventional PCR reactions consisted of the following: initial denaturation for 5 min at 95°C; followed by 35 cycles of 1 min at 95°C, 30 seconds at 59°C, and 1 min at 72°C; and a final extension of 7 min at 72°C. The PCR amplifications were conducted in volumes of 25 μl in a PTC-225 thermocycler (MJ Research, Watertown, MA, USA). The real-time PCR amplification was performed using a CFX96 real-time PCR system (Bio-Rad laboratories, Inc., USA) with the following cycling conditions: initial denaturation for 30 s at 95°C; then 45 cycles of 5 s at 95°C, 30 s at 59°C, and a melting curve of 65°C to 95°C at an increment of 0.5°C. The SYBR green real-time qPCR assay was performed in a 20-μl reaction. The TaqMan qPCR amplification of the Cms50, Cms72a and CelA primer sets was performed under previously described conditions (Gudmestad et al., 2009) (Table 2). The conventional and real-time PCR amplifications were carried out with the above primers (at a final concentration of 0.5 μM), GoTaq® DNA polymerase (1.25 μM final concentration; Promega, Madison, Wisconsin, USA) and SYBR® Premix Ex Taq™ (TAKARA BIO, Inc., Japan) according to the manufacturer’s instructions, using approximately 5 ng of purified DNA from each sample. After the conventional PCR reaction, 10 μl of the each PCR product was electrophoresed through a 1.5% agarose gel, stained with ethidium bromide, visualised on a UV transilluminator and imaged using a VersaDoc 1000 gel imaging system (Bio-Rad Laboratories, Inc., USA).
Specificity and sensitivity of the SYBR Green real-time qPCR assay
The specificity of the primer set Cms89FF/R was tested against a total of 12 strains of C. michiganensis subspecies and 22 bacterial strains from seven other genera. For sensitivity testing, the cloned DNA, total genomic DNA, and bacterial cell suspension from Cms NCPPB 2137 were serial diluted ten-fold and tested by SYBR Green real-time qPCR. The sensitivity evaluations were reproduced using serial dilutions and SYBR green real-time qPCR testing in triplicate. The data analysis, which included the determination of a standard curve, cycle threshold (Ct) or quantification cycle (Cq) value, PCR efficiencies, and slope of the standard curve, was automatically performed by the CFX ManagerTM Software system (Version 1.6; Bio-Rad Laboratories, Inc., USA). A copy number of the cloned DNA was calculated using the following equation:
Inoculation and sample preparation
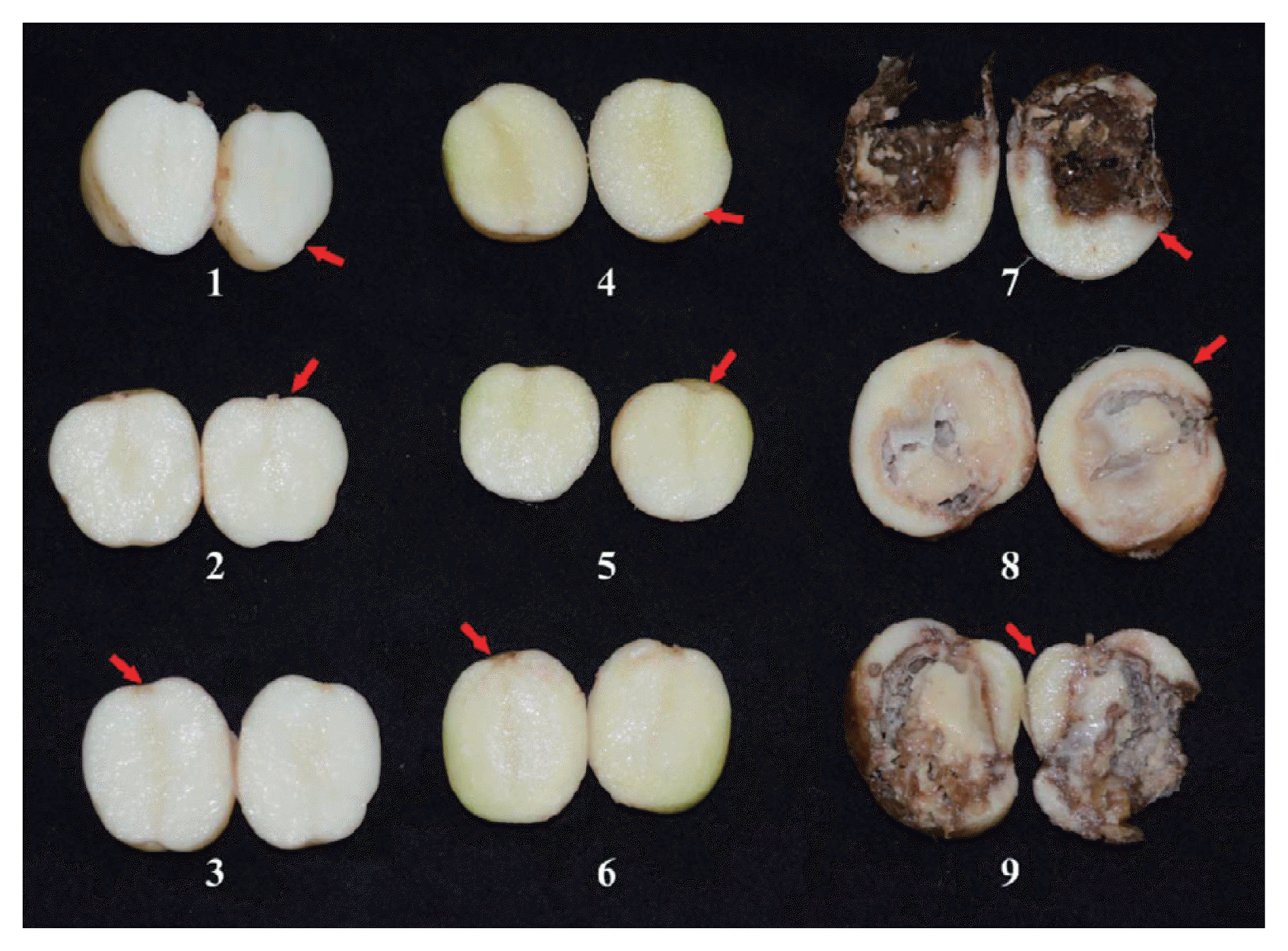
Cms-free potato tubers (Solanum tuberosum L. ‘Atlantic’) were removed from a cold storage facility one week prior to cutting and were inoculated as previously described (Slack et al., 1996). Briefly, the tubers were cut with one or two eyes and allowed to suberize for two days at room temperature. Then, the seed pieces were wounded by inserting a yellow pipette tip in both sides of the eye and then inoculating with 2 μl of a bacterial suspension (OD600nm = 2.0) containing either the NCPPB 2137 strain. The control seed pieces were inoculated with 2 μl of sterile distilled water. After inoculation, the seed pieces were allowed to rest for 30 min to stabilise the Cms. The pieces were then sown in 20-cm-diameter pots that were filled with an autoclaved artificial soil mixture; the sowed pieces were subsequently placed inside a greenhouse. The temperature inside of the greenhouse ranged from 20 to 25°C during the growing periods, and the tubers were watered daily until they reached the four-leaf stage. The experiment was carried out in a completely randomised blocked design with 15 tubers from five plants in each group and with each pot distant from the other pots to avoid cross-contamination. The 1-g samples were taken from an area of 1 cm that was distant from the symptom and symptomless lesion at 90 days post inoculation (dpi) as depicted in Fig. 1. Each sample was dipped in 500 μl of sterile distilled water for 30 min. The water that was treated with each sample (1 μl) was used in the SYBR green real-time bio-qPCR assays as described above.
Results
Bioinformatic analysis and primer design
Using the computational approach based on comparative genomics, a set of diagnostic primers was identified that distinguishes between three closely related pathogens: C. m. subsp. sepedonicus, C. m. subsp. michiganensis and C. m. subsp. nebraskensis (Lang et al., 2010). This approach also revealed some significant matches with previously determined sequences (BLASTN ver. 2.2.28+). BLASTN searches showed similarity to the conserved hypothetical protein sequence [identity=82%, Score=976 bits (528), Expect=0.0, and Gaps=8/1141] from C. m. subsp. nebraskensis NCPPB 2581. BLAST searches with the predicted protein sequence (BLASTX) revealed similarities with the conserved hypothetical protein sequence [identity=85%, Score=621 bits (1601), and Expect=0.0] from C. m. subsp. nebraskensis NCPPB 2581 and the hypothetical protein sequence [identity=33%, Score=149 bits (376), and Expect=9e-38] from Blastococcus saxobsidens DD2. However, the region to be amplified with the designed primer set revealed no significant match in either the BLASTN or BLASTX searches.
Specificity of the PCR assay
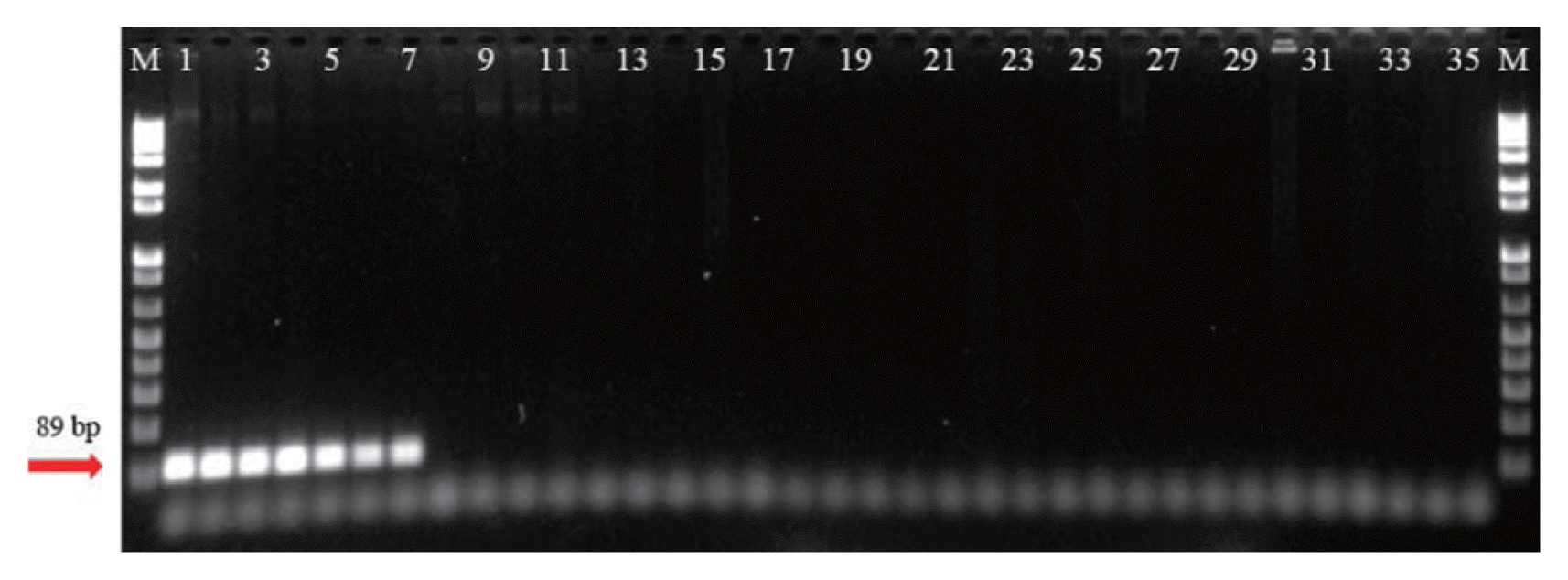
The specificity of the Cms89F/R primer sets was evaluated by conventional PCR and by the SYBR green real-time qPCR assay using reference strains (Table 1). Cms produced a positive band with the expected DNA fragment length (89 bp); however, the other Clavibacter subspecies strains and reference microorganisms did not produce any amplification product (Fig. 2). Only the assays with Cms yielded amplified products with fluorescence intensity and a single amplified DNA fragment. No signal was obtained from the negative control of the amplification reaction (NTC).
Sensitivity test by SYBR Green qPCR
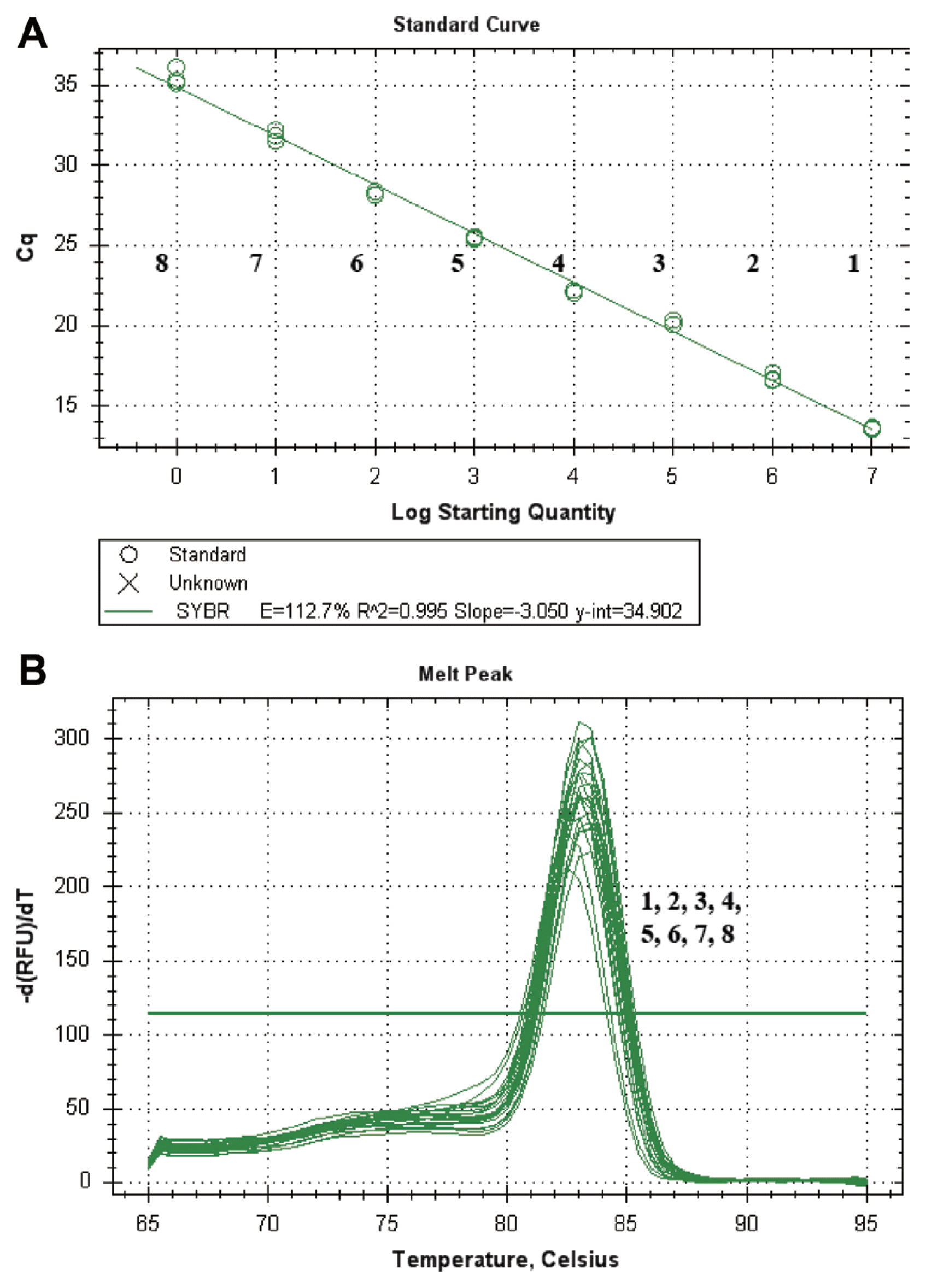
A standard curve that was based on cloned DNA was constructed to determine the Cms NCPPB 2137 concentration; a ten-fold serial dilution factor was used to achieve 1.47×109 gene copies per SYBR green qPCR. The amplification product of the targeted gene (putative phage-related protein) was cloned and used as control DNA for quantification. This assay was able to quantify at least 1.47 copies/μl of target DNA. The standard curve revealed a good linear response (R2=0.995). The standard regression analysis of the linear part of the slope produced a coefficient of −3.050, yielding a PCR efficiency of 112.7% (Fig. 3A). The analysis of the melting temperature and melting peaks of Cms by the SYBR green qPCR revealed a reproducible melting temperature of 83°C and specific peaks (Fig. 3B). The standard curves revealed a linear correlation between the Ct values and the concentrations of genomic DNA (R2 = 0.998, slope = −3.163) and bacterial cell suspension (R2 = 0.994, slope = −3.068) of Cms NCPPB 2137.
The limit of detection (LOD) was determined for the genomic DNA and bacterial cells as the probability of having a positive SYBR green real-time qPCR result greater than 95%. Measuring the proportion of positive replicates per known input amount of DNA, the LOD was established as 1.47×102 copies/μl of cloned-amplified target DNA, 5 fg of DNA using genomic DNA or 10−6 dilution point of 0.12 at OD600 units of cells per reaction (Table 3). Importantly, when present primer sets compared to the Cms50, Cms72a and celA gene-based primer sets, Cms89 primer sets were able to detect genomic DNA and bacterial cells at an at least ten-fold higher sensitivity than previous studied primers, respectively.
Detection of the pathogen by quantitative Bio-PCR in tuber infection
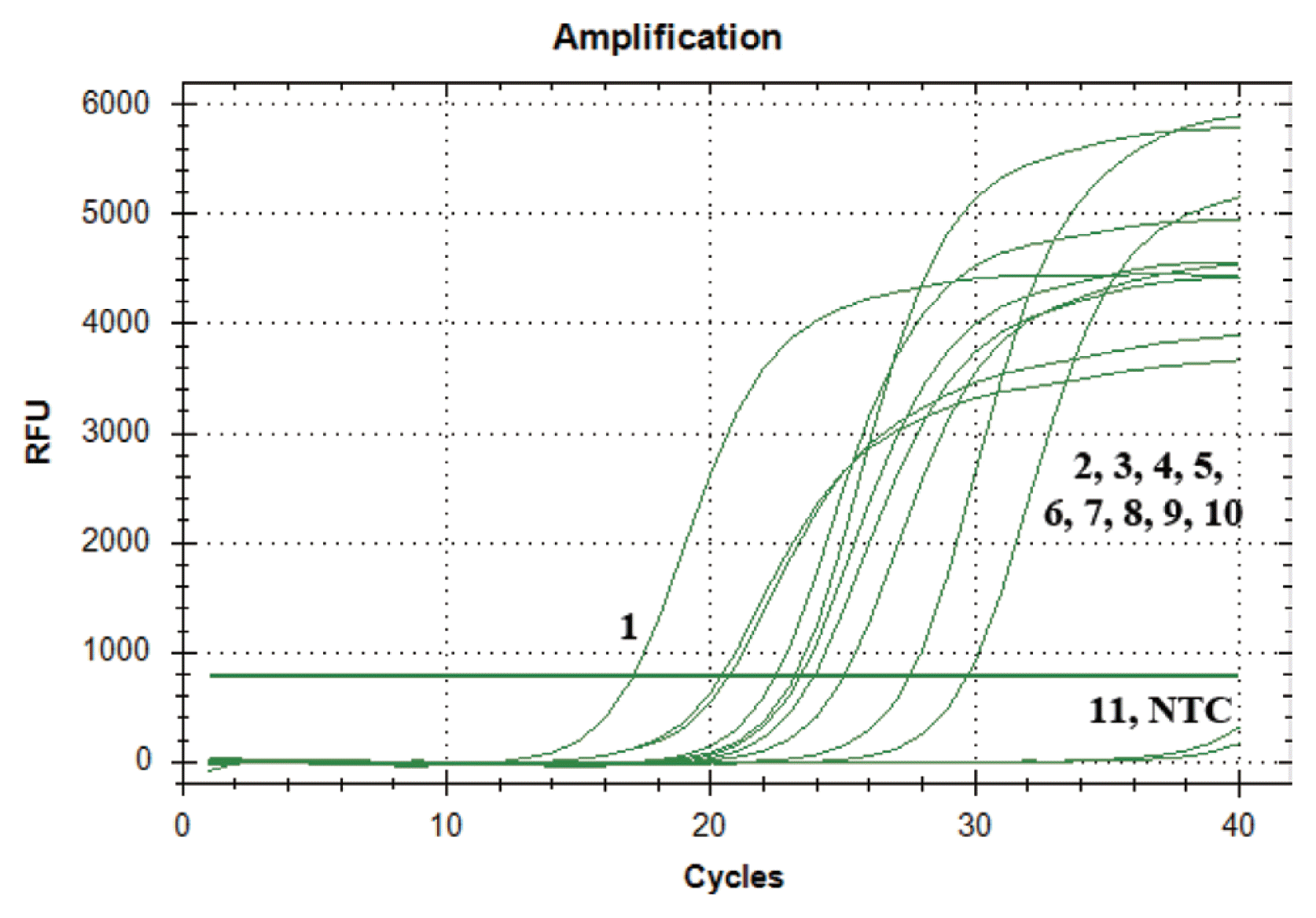
Using SYBR green real-time qPCR, the primer set Cms89F/R detected the pathogen in 15 of harvested potato progeny samples from inoculated artificially seed potato. The amplified products that were derived from the tubers both symptom and symptomless showed fluorescence in the SYBR green real-time qPCR assay throughout twice repeat experiments, while no signals were amplified from the healthy potato progeny samples (Fig. 5). The infected potatoes revealed Ct values ranging from 20 to 30.
Discussion
C. michiganensis subsp. sepedonicus is an emerging concern for international quarantine procedures and is an economically important disease for the potato seed industry worldwide. This pathogen can survive in potato tubers for long periods without producing symptoms (Pastrik, 2000). Therefore, sensitive detection and accurate identification are important for the prevention and management of this pathogen. Moreover, there is a high need for specific markers for the earlier detection and identification of Cms because currently available diagnoses result in large economic losses (Bentley et al., 2008; van der Wolf et al., 2005).
In general, the development of specific primers and DNA probes from the 16S rDNA region (Lee et al., 1997; Li and De Boer, 1995; Mills et al., 1997; Pastrik, 2000; Rademaker and Janse, 1994; Schaad et al., 1999; Schneider et al., 1993), the cellulase A (celA) gene and a small cryptic plasmid (pCS1) to detect Cms by PCR has been proposed (Gudmestad et al., 2009), including multiplex PCR (Pastrik, 2000). However, these assays fail to identify target pathogens and somewhat produce false positives because of critical defects in selecting genes from plasmids or hypothetical proteins (Gudmestad et al., 2009). Therefore, tests are generally invalid, and the designs need to be revised (Phillippy et al., 2007).
Recently, due to the large and increasing numbers of available bacterial sequences in databases, along with the already employed bioinformatics tools, more reliable, rapid and cost-effective methods are developed for bacterial identification from a broad range of samples (Albuquerque et al., 2009; Cho et al., 2011; Lang et al., 2010). This sequence information could be exploited to provide rapid and accurate diagnostics to identify target pathogens and to distinguish the pathogens from other closely related species (Phillippy et al., 2007; Slezak et al., 2003).
In this study, we found the specific sequence region using sequence alignment search methods and developed a primer set for the specific detection of the putative phage-related protein from the Cms ATCC 33113 genome sequence by accessing information in GenBank (Accession No. AM849034.1, gi|169155030:861988-863124). The putative phage-related protein sequence was analysed by BLAST searches and was found to be highly variable among C. michiganensis subspecies. Many researchers who study pathogen identification and the detection of food-borne pathogens have studied various bacteriophage (or phage)-based probes (Casjens and Molineux, 2012; Leiman and Shneider, 2012; Shabani et al., 2008; Singh et al., 2012; Singh et al., 2013; Smartt and Ripp, 2011; Smietana et al., 2011; Zourob and Ripp, 2010). These probes are thermally stable and sensitive, with a high specificity to bacterial cells (Nanduri et al., 2007). Bacterial cells have specific receptor molecules or carbohydrate (polysaccharide) sequences outside of the cell that serve as the primary mechanisms by which phages are identified (Golshahi et al., 2011). Therefore, in this study, a SYBR Green real-time qPCR assay for the detection of Cms based on the putative phage-related protein was developed. To overcome potential false-positive or -negative reactions from Cms strains, this assay was performed using collections from various geographical origins.
In conclusion, the sensitivity, simplicity and reproducibility of SYBR green real-time qPCR make it suitable as a screening method to detect and quantify Cms in potato samples, particularly for Cms outbreaks, allowing for more rapid monitoring. The methods that were used in this study are appropriate for analytical purposes with excellent limits of detection and quantification.

 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






