



Potato (Solanum tuberosum L.) is one of the most important vegetable crops in Korea and worldwide. However, its susceptibility to plant diseases including late blight caused by Phytophthora infestans makes its cultivation challenging (Chycoski and Punja, 1996; Desjardins et al., 1995; Giddings and Berg, 1919; Hahm et al., 1978; Park et al., 2011).
Late blight is one of the devastating diseases affecting Solanaceous crops like potato and tomato in most potato producing parts of Korea (Hahm et al., 1978; Park et al., 2011; Ryu et al., 2005) and worldwide. The worldwide economic damages of potatoes due to late blight are 5.0 billion dollars annually in 2004 (International Potato Center, 2004; Latijinhouwers et al., 2004). Late blight occurrence is favored by high humidity, dew, wet weather, and moderate temperatures (10-27°C) (Choi, 2007). When the environmental conditions are favorable for P. infestans, the disease can spread rapidly and defoliate entire fields within 3 weeks (Kirk et al., 2004). Late blight causes premature potato vine and tuber rots, and thereby results in poor yields. Generally, late blight affects folia parts of both potato and tomato along with potato tubers and tomato fruit (Novy et al., 2006). Potato late blight is easily transmitted by affected potato tubers. Planting non-certified potato seeds saved or obtained from neighbors increases the risk of incidence of late blight in the field.
In Korea, there are a total of 18 pesticides registered for the control of potato late blight, however, 20-95% of late blight strain show resistance to the fungicide, metalaxyl (Kim et al., 2000; Lee et al., 1994; Park et al., 2010; Zhang et al., 2005). In the 1980s, resistant potato late blight strain appeared in European fields, in the Netherlands and Switzerland, followed by the occurrence of metalaxyl-resistant strains in Ireland and Israel (Cohen and Reuveni, 1983; Dowly and O’sullivan, 1981; Fry et al., 1992). Therefore, eco-friendly environmental products are in high demand in the pesticide market. With the increase in a national income, the interest in health, and consumption of eco-friendly and organic agricultural products is rapidly increasing. Biological control has been recognized as a method that can replace synthetic pesticides in controlling various plant diseases (Chet and Inbar, 1994; Folman et al., 2004; Quimby et al., 2002). Moreover, various antagonistic microorganisms and naturally derived non-toxic substances are being used as biological pesticides.
With the rapid spread of organic farming, various methods for improving soil health are being explored, but there are very few competant organic materials available. Various beneficial microorganisms exist in an environment rich in organic matter, in the soil around the rhizosphere of plants (Yeo et al., 2009). In organic potato cultivation fields, diseases caused by soil transmitted pathogens such as late blight, common scab, and soft rot are frequent and cause severe economic damage. It is, therefore, necessary to develop eco-friendly plant disease control technologies. Late blight is one of the most destructive diseases in organic as well as conventional potato cultivation in Korea. The disease is known to destroy the entire leaves or foliar parts of the plant within 2 weeks of infection in organic fields (Ryu et al., 2005).
In recent years, the interest in biological control of plant pathogens has significantly increased due to the need for the introduction of environmentally friendly alternatives to minimize the intensive use of chemical pesticides (Ongena and Jacques, 2008; Quimby et al., 2002). Besides the antibiotic effect, many microbial strains also show growth-promoting properties on the plants. Among bacteria, the strains from genera Bacillus, Pseudomonas, Streptomyces, and Agrobacterium play an important role in biological control as soil and plant-associated microorganisms (Athukorala et al., 2009; Shim et al., 2015). Biocontrol agents are defined as the products consisting of living microorganisms used to combat plant pathogens. However, until the last decade, biocontrol products constituted only 1% of the sales of all agricultural chemicals on the global market (Fravel, 2005). For most biocontrol agents, the antagonistic ability selected under limited conditions in the laboratory does not exhibit antagonistic effects in the field as their anchoring power in the rhizosphere is low. However, the use of biological control agents and products is indispensable in organic farming (Lynch et al., 2012).
In many cases, microorganisms such as Trichoderma harzianum (Elad and Kapat, 1999; Wang et al., 2021; Yao et al., 2016), Bacillus subtilis (El-Naggar et al., 2016), and Burkholderia gladioli (Kang et al., 2013) were sufficient to suppress the late blight disease in the field. Bacterial bio-control agents from the genus Bacillus, primarily Bacillus velezensis, have effectively controlled the common potato disease caused by R. solani in several countries for a decade (Asaturova et al., 2021; Dinu et al. 2019; El-Naggar et al., 2016).
This study was conducted with the objective to identify Bacillus sp. AFB2-2 strain with optimal temperature. The study is also an attempt to estimate the antifungal ability of AFB2-2 strain against P. infestans and identified by molecular analysis. Moreover, we screened the strain for the presence of cyclic lipopeptide synthesis genes and evaluated its efficacy in controlling potato late blight in greenhouse condition and an organic potato field.
Materials and Methods
Plant pathogens
Seven plant pathogens potato fusarium wilt (Fusarium oxysporum, KACC 43207); potato gray mold (Botrytis cinerea, KACC 40963), potato black scurf (Rhizoctonia solani, KACC 40130), potato white mold (Sclerotinia sclreotiorum, KACC 41069), hot-pepper damping-off (Pythium ultimum, KACC 40705), potato late blight (Phytophthora infestans, KACC 43073), and hot-pepper anthracnose (Colletotrichum gloeosporioides, KACC 40003) were used in this study, and were kindly provided by the National Agricultural Microorganisms Resource Center (KACC). Seven plant pathogens, except for P. infestance, were cultured in PDA (20 g potato, 18 g dextrose, 18 g agar, 1,000 ml sterile water) medium, and the late blight fungus was grown on V8-juice agar (150 ml V8-juice, 850 ml distilled water, 3 g CaCO3, and 18 g agar). The culture conditions were incubated at 20-25°C for 7 days with due consideration of the optimal growth conditions of the pathogens. Actively growing mycelia were selected from the plates and subcultured in freshly prepared V8-juice or PDA media for further incubation at their respective optimal growth temperature.
Isolation and culture of antagonistic bacteria
To select strains with excellent inhibitory effect on potato late blight, soil samples were collected from a depth of 15 cm centered on the rhizosphere of potatoes with excellent growth at an organic potato cultivation farmhouse in Sillim-myeon, Wonju-si, Gangwon-do, in mid-June. To increase the selection frequency of the endospore-forming Bacillus, 1 g of the treated soil sample and 9 ml of physiological saline (0.85% NaCl) were added and treated in a water bath at 80°C for 60 min to lower the density of gram-negative bacteria. The heat-treated soil sample was then diluted to a concentration of 10−4 to 10−6 in a glass tube containing 9 ml of sterilized distilled water, spread on the tryptic soy agar (TSA) medium, and incubated at 28°C for 2 days. After culturing, a single colony was isolated and purified, suspended in a cryotube (2.0 ml) containing 20% glycerin and then stored in a −75°C cryofreezer.
In order to test the antifungal activity of 320 bacterial isolates isolated from organic potato cultivation soil, the PDA medium was autoclaved and cooled to 50°C. Then, pure cultured P. infestans mycelium and F. oxysporum conidia were added, respectively, and a culture medium was prepared in advance. Antifungal activity of the isolates was primarily selected by inoculating the culture medium containing the plant pathogens by using a sterilized toothpick to inoculate the single colony of the tested isolate.
Identification of Bacillus sp. AFB2-2
The morphological and physiological characterization of the AFB2-2 strain was based on the gram reaction, shape, motility, endospore formation, and growth at different temperatures as well as on the NaCl concentrations in the in vitro assays.
The cell morphology of the strain was observed under transmission electron microscope (LEO 912AB, ZEISS, Oberkochen, Germany) at ×10,000 magnification using cells grown for 2 days at 28°C on the TSA medium and prepared on Formvar-coated grids (Plano, Wetzlar, Germany) followed by staining with 3% uranyl acetate, as described by Hartmann et al. (2010).
The gram reaction of the strain was determined by non-staining methods using potassium hydroxide (KOH), as described by Buck (1982) and Halebian et al. (1981).
The isolates were identified with reference to their biochemical profiles of single-carbon source assimilation ability using the API 50CH/B Test Kit (BioMérieux, Marcy l’Etoile, France), following the manufacturer’s instructions. The Analytical Profile Index (API) test kit results were interpreted using the API database of the Apiweb software (version 4.0, BioMérieux).
For the thermotolerant screening, the AFB2-2 strain was grown in tryptic soy broth (TSB) medium at 10°C, 20°C, 25°C, 30°C, 35°C, 37°C, 40°C, 45°C, 48°C, 50°C, 52°C, 55°C, and 60°C in an RTS-8 Multi-channel Bioreactor (Biosan Co. Ltd., Riga, Lativia) for 25 h and the growth was recorded as optical density (OD) with noninvasive real-time cell concentration measurement at 600 nm.
The AFB2-2 strain was tested for its salt tolerance based on the relative changes in their growth as per a previously published protocol (Shim et al., 2015). For evaluation, the salt tolerance ability of the newly isolated strains was performed using a 100 μl aliquot of a preinoculum to inoculate 50 ml of TSB medium supplemented with different concentrations (0%, 1%, 2%, 2.5%, 3.0%, 5.5%, 6.0%, and 7.5%) of NaCl. After inoculation, the bacterial growth was evaluated using the RTS-8 Multi-channel Bioreactor (Biosan Co. Ltd.) at 37°C for 25 h. The absorbance of the culture was evaluated automatically at the 30-min interval at 600 nm in the RTS-8 Multi-channel Bioreactor. The highest OD value obtained at different NaCl concentrations was considered as the maximum salinity tolerance capacity.
In vitro inhibition assay for P. infestans on solid media
The candidate isolates AFB2-2 were screened for their antagonistic ability against seven plant pathogens, including P. infestans, in vitro by using the dual culture technique using the method described by Estrella et al. (2001) and Kerkeni et al. (2007). Therefore, the AFB2-2 isolate was cultured by streaking at 2 cm from the edge of a 9 cm diameter Petri dish containing the PDA medium. An 8-mm mycelial agar disk was uniformly cut from the leading edge of a 5-day-old culture of seven plant pathogens with an 8 mm diameter corker borer and deposited in an opposed position 2 cm away from the other set of the plate. For the untreated plates, an 8 mm mycelial agar disk of the pathogens only was placed at 2 cm from the edge of a 9 cm diameter Petri dish containing only fresh PDA medium.
Three plates were used as replicates for each treatment as well as for the control. Inoculated plates were incubated at 25 ± 1°C until the fungal growth of the control plates reached the edge of the Petri dishes. The reduction in the mycelial growth of the seven plant pathogens was calculated using the formula suggested by Hwang et al. (2006) and Shim et al. (2015), as follows:
, where MI = mycelial growth inhibition (%) of the pathogen, RC = radial growth of the pathogen in the control plates (cm), and RT = radial growth of the pathogen treated with antagonistic bacteria AFB2-2 in a dual culture plate (cm).
Identification based on 16S rRNA gene sequencing and phylogenetic analyses
The pure isolated AFB2-2 colony was inoculated in a 9 ml TSB medium and cultured for 24 h with continuous stirring at 28°C, after which the genomic DNA was extracted by using the modified bead beater phenol method. For molecular identification, the selected AFB2-2 strain was subjected to 16S rRNA gene sequence analyses. The AFB2-2 strain was grown in TSB and the genomic DNA was extracted (Saitou and Nei, 1987; Sambrook et al., 1989). The 16S rRNA gene was amplified by PCR using the forward primer 27F 5′-AGAGTTTGATCCTGGCTCAG-3′ and the reverse primer 1492R 5′-GGTTACCTTGTTACGACTT-3′. The amplification of 16S rRNA gene sequences was performed using the Thermal Cycler (Applied Biosystems TC1, Pittsburgh, PA, USA) with a 25 μl reaction volume. Each reaction mixture contained a 2 μl 10× buffer with magnesium chloride (MgCl2), 0.5 μl of 10 mM dNTPs, 2 μl of 10 pmol-PCR primers, 2 units of Taq DNA polymerase (TaKaRa Hot-Taq, Tokyo, Japan), and 2 μl of DNA template (50-100 ng), and the volume was increased to 25 μl with double distilled sterile water. PCR was performed for the internal transcribed spacer (ITS) region using the following steps: (1) 94°C for 5 min; (2) 30 cycles of 92°C for 60 s, 58°C for 60 s, and 72°C for 90 s; and (3) a final extension step at 72°C for 5 min. For the ITS DNA fragment sequencing, template DNA (20 μl) was directly prepared from the PCR products by purification using a column-based purification kit (Qiagen, Hilden, Germany). Sequencing was then performed with Chunlab Co. (Seoul, Korea). The obtained 16S rRNA gene sequences were compared with sequences previously deposited in the National Center for Biotechnology database (NCBI, https://blast.ncbi.nlm.hih.gov/Blast.cgi). A molecular phylogenetic tree based on the amino acid sequence of the 16S rRNA gene of the AFB2-2 strain was performed automatically using the MEGA version 7 (Kumar et al., 2016) after multiple alignments of the data with the CLUSTAL W (Thompson et al., 1994). Distances were obtained using options according to the Kimura two-parameter model (Kim et al., 2016; Kimura, 1980), and clustering was performed using the Neighbor-Join algorithms (Saitou and Nei, 1987) to a pairwise matrix, which was inferred by using the maximum composite likelihood method, with a 1,000-replicate (Felsenstein, 1985) bootstrap test using MEGA 7. The phylogenetic tree was drawn to scale, with the branch lengths measured by the number of substitutions per site.
Screening of secondary metabolite biosynthesis genes of AFB2-2 strain
PCR was performed with specific primers to detect bacillomycin D (bmyA), fengycin (fenD), iturin A (ituA), surfactin (srfA), and zwittermicin A (zwiA) synthesis-related genes exhibiting antagonistic activity against Bacillus sp. strains (Table 1). For the PCR reaction, a reaction mixture (AccuPower HotStart PCR PreMix, Bioneer Co., Daejeon, Korea) was used to which 1 μl of genomic DNA (50 ng) and 10 pmol of each primer were mixed, with 20 μl set as the final volume. For the PCR reaction, the initial denaturation reaction (initial denaturation; 95°C, 3 min), amplification reaction (denaturation: 95°C for 30 s, binding: 55°C for 45 s, and elongation: 72°C for 1 min) was repeated 30 times, and the final elongation reaction was performed at 72°C for 10 min. The presence and size of the amplified PCR product were confirmed via electrophoresis on 1.2% agarose gel in 0.8% TSB buffer.
Control efficacy of AFB2-2 strain against potato late blight in the pot and field trials
Mycelium fragments of potato late blight (P. infestans) were placed on the V8-juice agar medium and cultured for 7 days in a low-temperature incubator under dark at 25°C. When the aerial mycelium on the surface was removed and placed under fluorescent light for 1 day, sporangia were formed. The sporangia were then harvested from the medium to form zoospores from sporangia, which were left in the refrigerator at 4°C for 2 h. The least population density of zoospore was more than 106-7 cells/ml that was drenched in the soil and spray on the 40-day-old Sumi potato plant at 2 h after treatment of 100 times diluted antifungal bacterial culture solution of AFB 2-2 in distilled water. 100 ml of each treatment was irrigated in the root and sprayed on the stems and leaves of potato plant. And the control treatment treated with spray of the 100 times diluted TSB media in distilled water. All experiments were conducted at least twice, a set of five replicates with 10 plants was performed per treatment. The number of infested plant was recorded every 7 days for 28 days after treatment.
To investigate the control effect of the AFB2-2 strain on potato late blight in organic potato cultivation fields in 2019. The potato of the Sumi cultivar was sown in a farmhouse field in Sillim-myeon, Wonju-si, Gangwon-do, on May 15, and then grown using the organic cultivation method. Field trials were performed according to Lamsal et al. (2013). A total of four treatments included control was set up for the experiment, with randomized complete blocks. In the farm field, 30 healthy potato seeds were sown at intervals of 30 × 70 cm in 6.6 m2, and the experiments was conducted with four repetitions of the completely randomized design for a total of 16 plots. A total of three investigations were conducted. From June 26, the beginning of the onset, the AFB2-2 culture solution was sprayed four times with 14-day intervals to sufficiently wet the aboveground portions, and then the late blight disease severity (%) was recorded up to 90 days of planting by the scale from 0 to 4 according to Cohen et al. (1991) based on the infected leaf area as follows: 0, no infested leaves; 1, 0.1-25% infested leaves area; 2, 25.1-50% infested leaves area; 3, 50.1-75% infested leaves area; 4, more than 75.1% infested leaves area.
Percentage of infected plant (%) was recorded up to 90 days of planting and based on the infected leaf area as follows:
Weather conditions
The meteorological parameters (soil and atomospheric temperature) were recorded by using a portable weather station WatchDog 1000 Series Micro Stations (Spectrum Technologies Instruments, Aurora, IL, USA) at 1.2 m height inside the study field. The soil temperature was detected by SMEC 300 soil moisture/EC/Temperature senor (Spectrum Technologies Instruments) placed in 10 cm depth of soil under black PVC film. The records were extracted per hour.
Statistical analysis
The step of gathering the experimental data was repeated at least thrice. All the data were subjected to statistical analysis using the SAS (version 9.2_PC32, SAS Institute Inc., Cary, NC, USA). Significance testing between treatments was analyzed using Duncan’s multiple range tests at a 5% level.
Results
Evaluation of biological properties of AFB2-2 strain
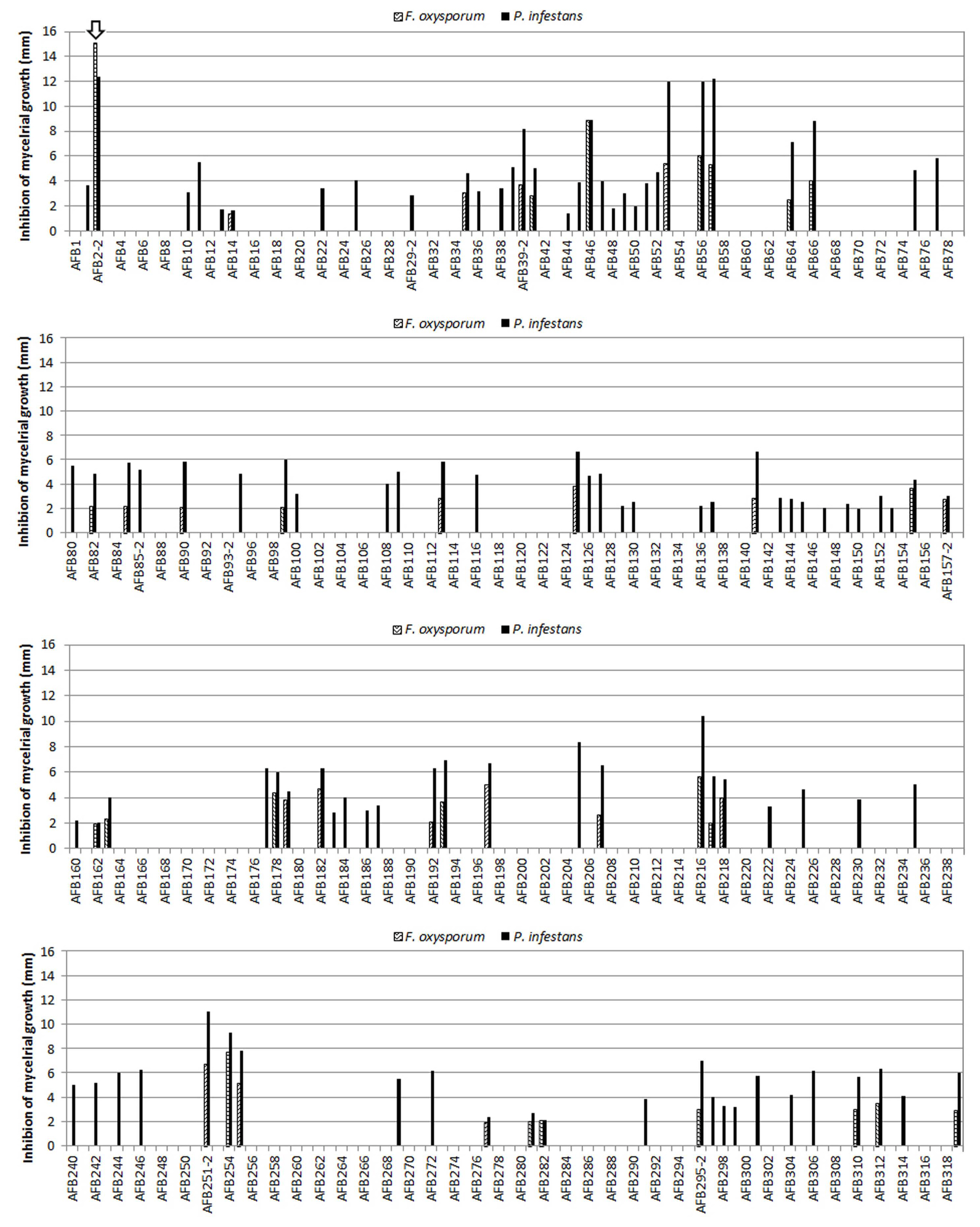
Among a total of 320 isolates from organic potato cultivation soil, the AFB2-2 strain, showing the highest mycelial inhibition in P. infestans, was identified (Fig. 1). AFB2-2 strain is a gram-positive spore-forming motile bacterium with peritrichous flagella. Observation under the transmission electron microscope revealed that its cell size was (1.56 ± 0.13) × (0.58 ± 0.05) μm (Table 2, Fig. 2).
Evaluation of carbon utilization ability of AFB2-2 strain
The utilization of 49 carbon sources by AFB2-2 strain was tested using API 50CH Kit at two incubation temperatures 28°C and 35°C. At 28°C, the AFB2-2 strain was able to utilize 11 carbon sources glycerol, D-glucose, D-fructose, D-mannose, D-mannitol, D-morbitol, esculin ferric citrate, D-cellobiose, S-saccharose, D-trehalose, and gentiobiose (Table 3). At 35°C, AFB2-2 strain utilized 20 kinds of carbon sources, glycerol, L-arabinose, D-ribose, D-glucose, D-fructose, D-mannose, inositol, D-mannitol, D-sorbitol, esculin ferric citrate, salicin, D-cellobiose, D-maltose, D-lactose, S-saccharose, D-trehalose, D-raffinose, amidon (starch), glycogen, and gentiobiose (Table 2). At 35°C, which is the near-optimal temperature for the growth of bacterial cultures, the AFB2-2 strain was found to utilize nine more carbon sources than that at 28°C (Table 2).
In Table 3, the biochemical test results of API50CHB of the AFB2-2 strain were compared with the previously reported biochemical test results of the Bacillus group, showing a higher similarity with the B. velezensis group in the use of L-arabinose, D-mannose, and D-trehalose.
Evaluation of temperature and NaCl tolerance ranges for AFB2-2 strain
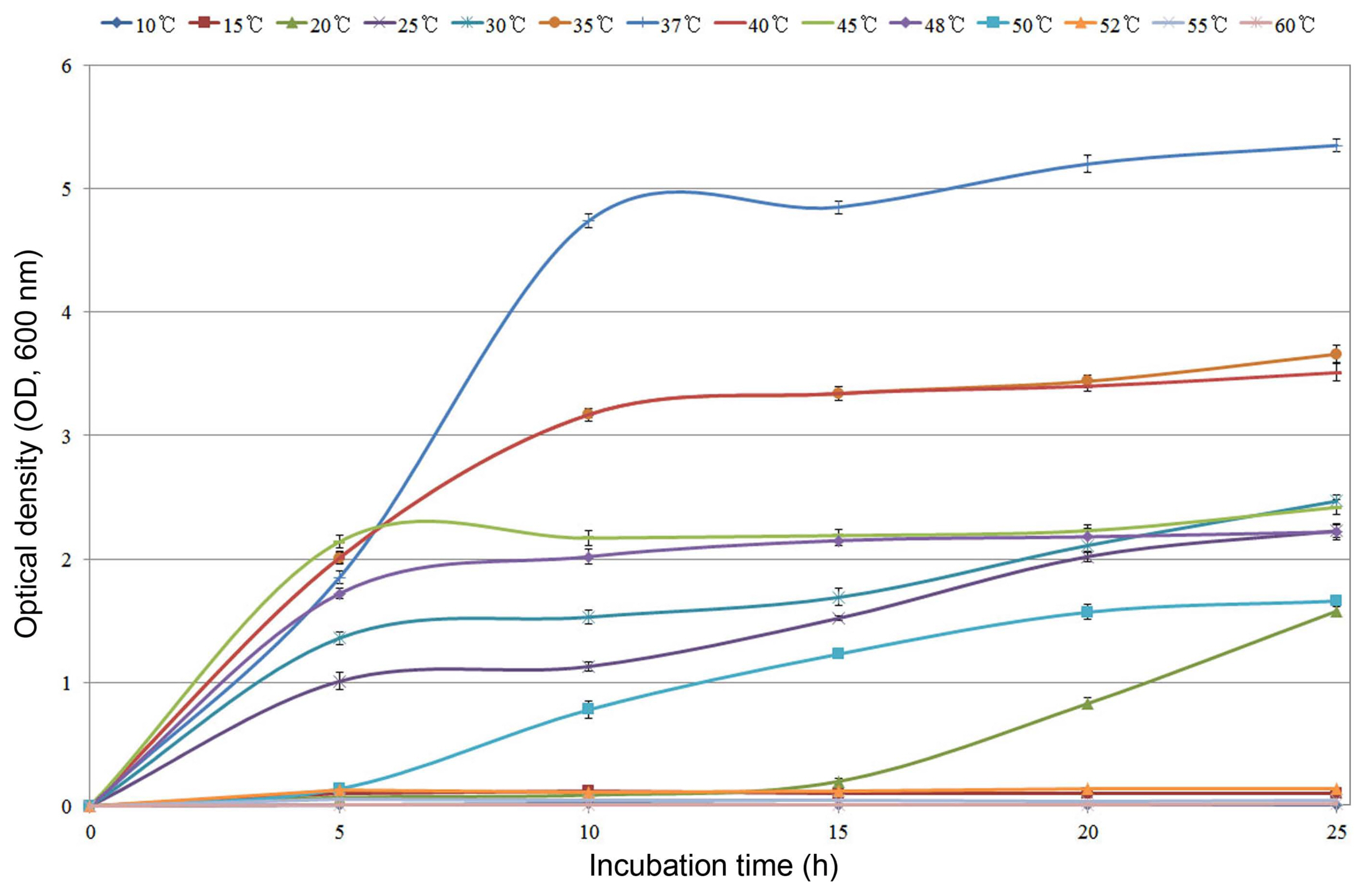
AFB2-2 strain was evaluated for their ability to grow at a wide range of temperature conditions (10-60°C) in the TSB medium. The strain showed normal growth in the range of 20-50°C; however, higher temperatures (52-60°C) hampered its growth. It showed optimum growth at 37°C with an OD600 of 5.5 at 25 h after inoculation in the TSB medium (Fig. 3).
A step-by-step estimation of NaCl tolerance revealed that the AFB2-2 strain could tolerate up to 7.5% of NaCl. The AFB2-2 strain was cultured at different salt concentrations, like 0%, 1%, 2%, 2.5%, 3%, 5.5%, 6%, and 7.5% of NaCl, in the TSB medium to study its salt requirement (Table 4). In the TSB liquid medium, 7.5% NaCl is a critical point for the growth of the AFB2-2 strain. AFB2-2 strain showed optimal growth at 2.5% NaCl, with OD 600 of 2.64 at 25 h after inoculation (Table 4).
Estimation of antifungal activity test of AFB2-2 strain against plant pathogens
The growth inhibitory effect of AFB2-2 strain was tested on the mycelia of seven major plant pathogens, F. oxysporum, B. cinerea, R. solani, S. sclreotiorum, P. ultimum, P. infestans, and C. gloeosporioiedes. AFB2-2 strain inhibited the mycelial growth of seven of these plant pathogens with an average zone of 10.2 mm or more, 7 days after inoculation (Table 5). Among the seven plant pathogens, P. infestans, and C. gloeosporioiedes showed the highest mycelial growth inhibition compared with the untreated control at 73.0% and 71.2%, respectively. In particular, the mycelial growth inhibition by 12.4 mm was observed against P. infestans (Table 5). The AFB2-2 strain showed a mycelial growth inhibitory effect of 40.3% or more compared with the untreated plant pathogens cultures (Table 5).
Control efficacy of AFB2-2 strain against potato late blight in greenhouse conditions
The efficacy of the AFB2-2 strain in controlling the potato late blight caused by P. infestans was assessed in the greenhouse. A 5-day-old culture solution of the AFB2-2 strain was diluted to 105, 106, and 107 cfu/ml and sprayed on 50-day old potato plants followed by the treatment with P. infestans zoospore suspension (106-7 cells/ml). The plants were examined after every 7 days after spray inoculation till the 28th day. The disease incidence of potato late blight in untreated potatoes was 46.7% at 7 days after inoculation and 93.3% on the 28th day (Table 6). At 7 days after the spray treatment, the AFB2-2 treatment showed a difference in disease incidence with an average of 17.9% or less. Among the three concentrations of AFB2-2 treatment, the 107 cfu/ml treatments exhibited a low rate of disease incidence at 2.2% and a high control value at 95.2% (Table 6). After 28 days of treatment, disease occurrence was up to 93.3% in untreated potatoes but 33.3% or less on average in AFB2-2 treated potatoes. The occurrence of potato late blight was suppressed by AFB2-2 treatment from 7 days to 28 days after treatment. Among the three AFB2-2 treatment concentrations, the control value of 107 cfu/ml of AFB2-2 treatment was the highest at 85.7% (Table 6).
Control efficacy of AFB2-2 strain against potato late blight in the farm field
To further test the effectiveness of the AFB2-2 strain, naturally occurring potato late blight caused by P. infestans was subjected to ABF2-2 treatment. A typical field outbreak of potato blight in Sillimmyeon, Gangwon-do in 2019 was used for this evaluation. Thus, when the incidence rate was 2% or more on average, ABF2-2 culture solution diluted to 105, 106, and 107 cfu/ml was sprayed. The inhibitory effect was investigated till the 42nd day at 7 day intervals.
In the case of untreated control, after 7 days of treatment, the potato late blight incidence rate was 24.2%. It increased over time to 42.3% on the 42nd day, when the disease severity was 0.76 (Table 7). The disease incidence rate of potato late blight in AFB2-2 treated potatoes differed depending on the treatment concentrations, but the average was 8.9% or less (Table 7). The occurrence of disease in untreated potatoes was high (42.2%) 42 days after AFB2-2 treatment, but in the case of AFB2-2 treated potatoes, it occurred at an average of 13.3% or less. Among the three concentrations of AFB2-2 treatment, the disease incidence and severity in the 107 cfu/ml treatment was lowest at 0.07 and 6.7, respectively, whereas the control value against on the 42nd day was the highest at 84.2% (Table 7).
Monitoring of air and soil temperature in the field
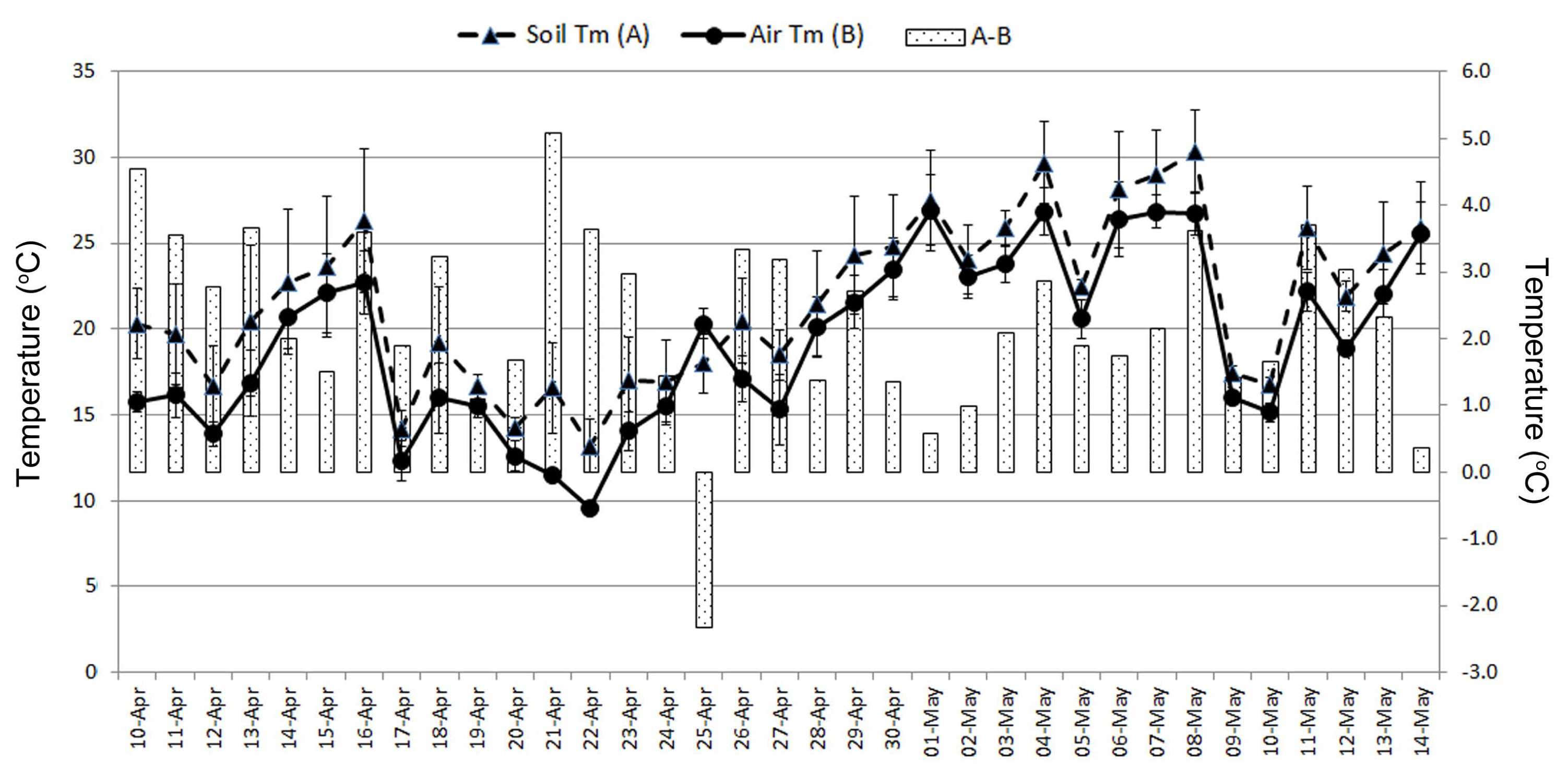
During the test period, the air and soil temperature of organic potato cultivation field were continuously monitored at Wonju from April 10 to May 14, 2020. The daily (24 h) maximum air and soil temperatures are nominally recorded at from 10 am to 2 pm. Air temperatures during the cultivation of organic potato were between 8.7 and 29.5°C for all experiments. And, soil temperatures of the field were 9.9 to 32.7°C (Fig. 4). Between April 30 and May 8, the air and soil temperature of the potato field was maintained above 25°C. Moreover, during the test period, except for April 25, most of the soil temperature was higher than the atmospheric temperature by 0.4°C or more (Fig. 4).
Identification of AFB2-2 strain based on biological and molecular characteristics
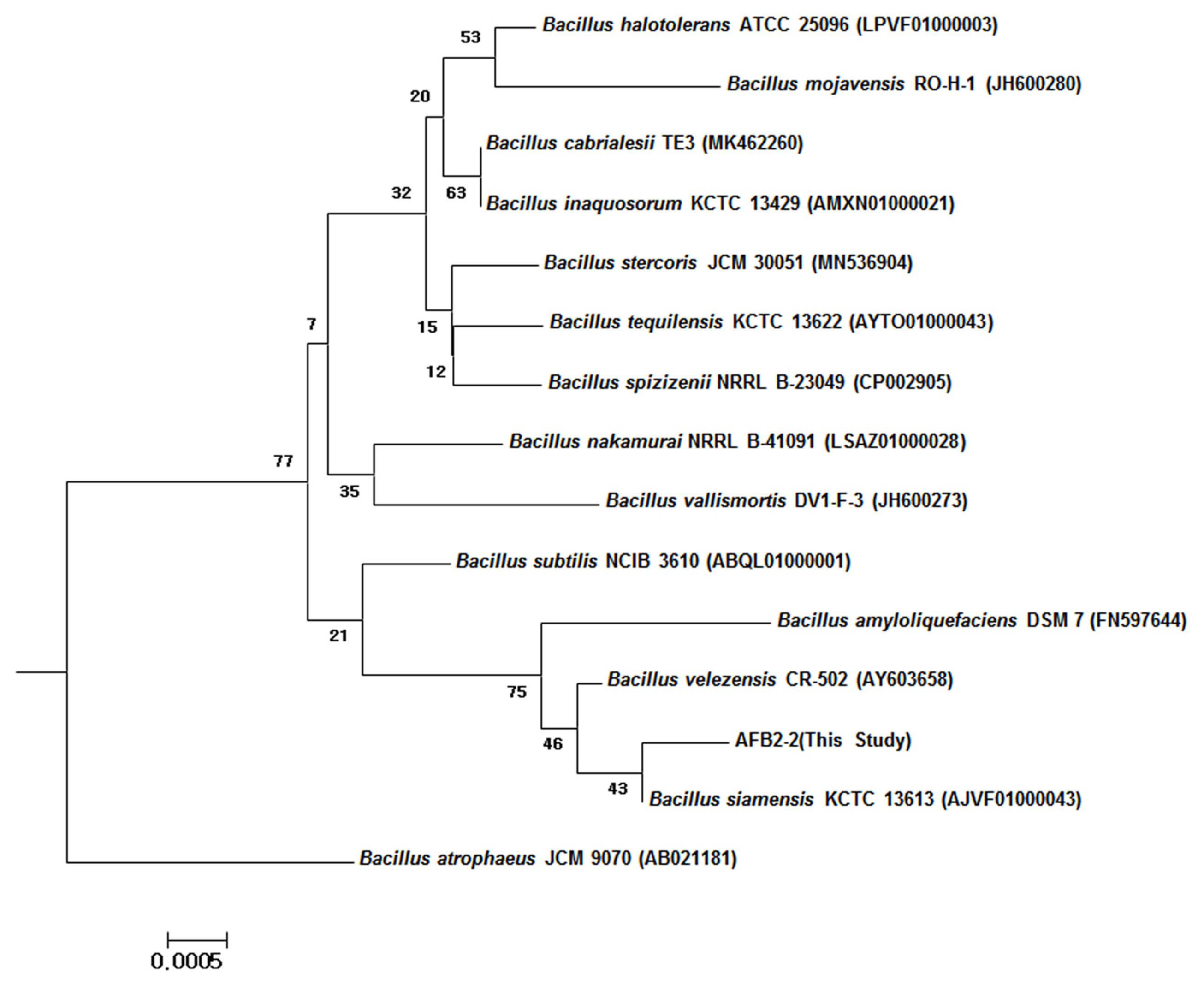
The isolated strain AFB2-2 had a rough and wrinkled white color on the surface of the colony, and the form was a gram-positive bacterium of Bacillus in Table 2. As a result of identification of the strain through the study of sugar availability using the API 50 CHB system (Table 2), it was identified as Bacillus velezensis (synonym of B. amyloliquefaciens) (data not shown). For more accurate identification, after determining the 16S rRNA gene sequence of the isolated strain (1.5 kb) was amplified from purified genomic DNA of the AFB2-2 strain. The amplified nucleotide sequence of 1,492 bp was sequenced and analyzed. The obtained sequences were searched against the NCBI GenBank database. Bacillus siamensis strain KCTC 13613, Bacillus velezensis strain CR-502, and Bacillus amyloliquefaciens strain DSM7 were found to have a similarity of 99.7%, 99.7%, and 99.5%, respectively (Fig. 5). As shown, the isolated strain was named B. velezensis AFB2-2, and the base sequence of the 16S rRNA gene was registered in GenBank.
Detection of secondary metabolite biosynthesis genes of B. velezensis AFB2-2 strain
B. velezensis AFB2-2 strain showed prominent antibacterial activity against various plant pathogens. PCR-based detection of biosynthetic genes revealed that it harbors three secondary metabolite biosynthesis genes, namely, bacillomycin D (bmyA, 875 bp), iturin A (ituA, 647 bp), and surfactin (srfA, 441 bp) (Fig. 6). However, genes related to fengycin and zwittermicin production were not detected (Fig. 6).
Discussion
Recently, interest in organic agriculture has been growing considerably as the demand for environmentally safe agricultural products has rapidly increased. However, research on eco-friendly agricultural technologies is needed to prevent the damage from pests and diseases for increasing the yield.
The incidence and progress of the disease largely depend on climatic conditions around the fields (Miller et al., 1998). Potato leaves in conventional farming fields remain healthy for 5 weeks after the appearance of the first symptom, as fungicides are applied four times on an average to control the disease (Ryu et al., 2005). However, potato late blight is not easy to control and manage by organic/biological means. Organic farmers should employ multiple strategies available to reduce late blight risk in host crops (Ryu et al., 2005).
The morphological and biological characteristics of AFB2-2, characterized in the present work, were similar to the B. velezensis described by Kim et al. (2017) and Chen et al. (2018). Our morphological identification was confirmed by amplification and sequencing of a partial fragment of the 16S rRNA region (Smibert and Krieg, 1994).
Recently, Chun et al. (2019) and Jin et al. (2018), L-arabinose, D-xylose, and D-mannose in a study comparing the biochemical properties of the B. amyloliqefaciens group and the B. siamensis group similar to the B. velezensis group. And it was reported that there is a difference in the use of D-trehalose. In particular, it suggests that the difference can be distinguished from the B. amyloliqefaciens group, which shows the most similarity in 16S rRNA analysis.
In this study, as a result of 16S rRNA analysis of the AFB 2-2 strain, it was found that the B. siamensis group and the B. amyloliqefaciens group were closest to each other. However, the assay results for biochemical availability using the API50CHB were reported by Chun et al. (2019) and Jin et al. (2018), it is judged to show higher similarity to the Bacillus velezensis group in the use of L-arabinose, D-xylose, D-mannose and D-trehalose. Therefore, we identified the AFB 2-2 strain as B. velezensis AFB 2-2 based on the overall results from the morphological, biochemical test and molecular data.
Bacillus, Pseudomonas, and other microorganisms have been extensively studied for their ability to solubilize nutrients, biocontrol potential, and plant growth-promoting abilities in all crop production systems. However, successful deployment of these organisms in ecosystems under stress depends on their ability to withstand and proliferate under adverse environments such as heat, salinity, heavy metal toxicity, and so forth (Praveen Kumar et al., 2014; Lucy et al., 2004).
Several studies have emphasized the use of beneficial microorganisms, which can tolerate heat stress and augment the growth and productivity of crop plants (Abd El-Daim et al., 2014; Ali et al., 2009, 2011; Khan et al., 2020; Park et al, 2017; Srivastava et al., 2008). In this study, B. velezensis AFB2-2 was a strain capable of growing at a high temperature of 50°C, and its stable culture was possible at 37°C in a liquid medium.
Salt stress tolerance is an important property of saprophytes and competitiveness among rhizobial isolates. Therefore, it was observed that some of the salt-tolerant isolates might have good saprophytic and competitive abilities to perform well in the rhizosphere of soil (Praveen Kumar et al., 2014). The salt-tolerant microbes were domesticated to tolerate higher concentrations of NaCl, which can play a biological role in high salt stress in field conditions.
Salt-tolerant bacteria can enhance the ability of plants to withstand salt stress. The bacterium has a special cell membrane and cell wall structure, which can effectively prevent Na+ from entering the cell, and maintain a low salt concentration in the cell. Thus, it can play a greater role in promoting the growth and the cultivation of salt-tolerant plants (Hussein and Joo, 2018; Masmoudi et al., 2019; Siddikee et al., 2010; Wang et al., 2021).
Soil salt accumulation has been steadily increasing over the past 10 years and is pointed out as a major inhibitor to crop growth along with soil heavy metals and pathogens (Hussein and Joo, 2018). The AFB2-2 strain could be cultured at a concentration of 7.5% of NaCl and it might potentially survive in soil with a high salt concentration.
Potato (Solanum tuberosum L.) is the third most important food crop consumed in the world after rice and wheat. Late blight, caused by the oomycete pathogen P. infestans, is considered the most devastating disease in the cultivation of potato (Kamoun et al., 2015).
In total, 149 types of microbial pesticides have been registered worldwide and used against pests and diseases. Bacillus subtilis QST 713 (RhapsodyR, SerenadeR) and Bacillus pumilus QST 2808 (Sonata R) have been developed for disease control and used as eco-friendly biological pesticides in Korea and many other countries (Cawoy et al., 2011; Copping, 2004).
In this study, B. velezensis (AFB2-2 strain) could suppress potato late blight caused by P. infestans in the greenhouse and natural field conditions. The B. velezensis (AFB2-2 strain) showed substantial growth inhibitory effect against the mycelia of seven important plant pathogens including P. infestans, and possesses three secondary metabolite biosynthesis genes namely, bacillomycin D, iturin A, and surfactin.
Secondary metabolites produced by Bacillus strains, related to plant disease control are cyclic lipopeptides (bacillomycin D, fengycin, iturin, surfactin), siderophore (bacillibactin), polyketides (bacillaene, difficidin, and macrolactin), a dipeptide (basilysin), acetoin, 2, 3-butandiol, etc. (Lee et al., 2012; Ramarathnam et al., 2007). These can aid in the inhibition of mycelial growth or spore germination in fungal pathogens, induction of resistance, and formation of biofilms to prevent plant diseases. It can be used in farms as an eco-friendly control strategy in organic potato cultivation.
Bacillus strains produce various secondary metabolites owing to the presence of biosynthetic genes. Genes for synthesis of bacillomycin D and fengycin were found to be present in the GH1-13 strain, which produced the relevant antibiotics (Kim et al., 2016). Kim et al. (2012) reported that Bacillus amyloliquifaciens GR4-5 strain harbored the fengycin biosynthesis gene in addition to the bacilycin biosynthesis gene, and their antibacterial activity was also confirmed. Moreover, Kim et al. (2016) reported that the GH1-13 strain possessed four types of genes: bacillomycin D (875 bp), bacilycin (749 bp), fengycin (964 bp), iturin (647 bp), and surfactin (441 bp). PCR was performed using gene-specific primers for detecting, ituA (iturin A), srfA (surfactin), and zwiA (zwittermicin A).
In conclusion, although potato is one of the most important vegetable crops in Korea and across the world, its susceptibility to diseases like late blight caused by Phytophthora infestans makes its cultivation challenging. In our study, Bacillus velezensis AFB2-2 strain, which has excellent antifungal activity against major plant pathogens, was characterized for its mycelial growth inhibition and identified by molecular analysis. This strain was also screened for the presence of cyclic lipopeptide synthesis genes and evaluated for its efficacy in controlling potato late blight in an organic potato field. We found that the AFB2-2 strain is a gram-positive spore-forming bacterium with peritrichous flagella and motility that can utilize 20 types of carbon sources, with the optimum growth temperature at 37°C. Its maximum salt concentration tolerance is 7.5% NaCl. This strain could inhibit the mycelial growth of seven plant pathogens after 7 days. After 42 days of AFB2-2 treatment, the disease occurrence in AFB2-2 treated potatoes was 13.3% or less. This strain was found to harbor the biosynthetic genes for bacillomycin D, iturin, and surfactin. We believe that the present findings on the characterization of the AFB2-2 strain can be used to develop an effective microbial agent for inhibiting the growth of fungal and bacterial pathogens against various crops through organic agriculture.
Although the late blight disease is an important fact in eco-friendly potato cultivation in Korea. Potato late blight resulted in annual yield losses of several billion dollars, in addition to the negative effects on the environment of frequent use of fungicides. For these issues, environmental products increasingly appear to be a significant portion of the pesticide market. Recently, it is highly dependent on the use of eco-friendly agricultural materials and the developments of biological control results are low. Biological control is an alternative technique to synthetic chemical fungicides in control of plant diseases, in order to create a more sustainable agricultural system


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






