



To ensure food security in the foreseeable coming decades, agricultural production must boost to keep pace with the expanding human population. There is currently a tremendous drive to create low-input and more sustainable agricultural strategies that include replacements to chemicals for pest and disease management. Thus, it is crucial to investigate the function of soil microorganisms which can stimulate growth and prohibit disease, as well as their use as biological control agent (Ab Rahman et al., 2018).
The Fusarium genus is one of the most sophisticated and versatile in the Eumycota, and the Fusarium oxysporum species group features plant, animal, and human diseases, as well as a varied diversity of non-pathogens (Gordon, 2017). Members of the Fusarium genus are a widespread soil-borne pathogen that infects a broad variety of horticultural and food crops, causing devastating vascular wilts, rots, and damping-off diseases (Bodah, 2017). F. oxysporum penetrates the root epidermis, spreads through the vascular tissue, and conquers the plant’s xylem vessels, resulting in vessel blockage and severe water stress (Singh et al., 2017). The tomato (Lycopersicon esculentum) is a crop grown on a global scale and is eaten raw or processed in a variety of products (Brookie et al., 2018; Srinivas et al., 2019) and its production is largely hampered by Fusarium wilt caused by F. oxysporum f. sp. lycopersici (Borisade et al., 2017).
Dickeya chrisanthemi (Dc, formerly Erwinia chrysanthemi) is an enterobacteria that causes blackleg and soft rot diseases on a wide variety of agricultural and aesthetic plants, most notably banana, Chrysanthemum spp., Dianthus spp., maize, potato, and tomato (Ma et al., 2007; Toth et al., 2011). The signs of soft rot are mostly produced by secreted pectinases that breakdown pectin in the intermediate lamella and primary cell walls (Hugouvieux-Cotte-Pattat et al., 2014). Dc is a serious bacterial pathogen associated with potato soft rot disease (Cating et al., 2008; Pérombelon, 2002; Potrykus et al., 2016; Samson et al., 2005; Sławiak et al., 2009; Toth et al., 2011; Van der Wolf et al., 2014; Végh et al., 2014).
Rhizosphere-resident fungi that benefit plants are often referred to as ‘Plant growth promoting fungi’ (PGPF). They are also a major source of biotic inducers known to confer several benefits on their host plants, and hence play a critical role in sustainable agriculture (Murali et al., 2021). The well-known fungal genera Aspergillus, Fusarium, Penicillium, Piriformospora, Phoma, and Trichoderma are the most commonly reported PGPF when it comes to ensuring improved plant development and induced systematic resistance (Hossain et al., 2017). Current findings have shown that Aspergillus spp. isolated from soil increases the synthesis of indole-3-acetic acid (Abri et al., 2015; Kumar et al., 2017; Ozimek et al., 2018), phosphorus solubilization (Li et al., 2016), and zinc oxide solubilization (Šebesta et al., 2020). Siderophore formation (Ghosh et al., 2017; Mahmoud and Abd-Alla, 2001; Nenwani et al., 2010; Patel et al., 2017; Salas-Marina et al., 2011), starch hydrolysis (Tsukagoshi et al., 2001) and antimicrobial secondary metabolites production (Lenartowicz et al., 2015). Secondary metabolites are low molecular weight molecules generated by fungi that are not essential for growth or reproduction but impose a competitive advantage on the organism. For instance, they may limit the development of organisms with which they interact, so impairing often vital biochemical reactions (Mosunova et al., 2020). The genus Aspergillus is the most prevalent, yielding an abundance of beneficial bioactive antimicrobial secondary metabolites (El-hawary et al., 2020; Ortega et al., 2021).
Aspergillus terreus is a significant saprophytic filamentous fungus found in soil that performs a variety of growth-promoting roles and produces antimicrobial secondary metabolites such as phenols and flavanoids (Javed et al., 2020; Vassileva et al., 2020). Additionally, some studies indicate that it has antifungal (Abdallah et al., 2015; Al-Shibli et al., 2019; Joncy et al., 2019; Waqas et al., 2015) and antibacterial properties (Cazar et al., 2005; Liu et al., 2018; Zaman et al., 2020). Additional research is required to determine its potential not only as a growth promoter for plants, but also as a biocontrol agent against a number of vital plant diseases.
The objective of the present study is to isolate fungal strain with biocontrol activities from the soil. ANU-301 isolated from the screening was identified as A. terreus by morphological and molecular analysis. Importantly, ANU-301 significantly suppressed tomato Fusarium wilt and potato soft rot. To our knowledge, it is the first report of biological control activity of A. terreus against F. oxysporum and Dc.
Materials and Methods
Plant pathogens
Fungal (Colletotrichum acutatum, Alternaria alternata, and F. oxysporum) and bacterial (Dc) plant pathogens were used in this study. F. oxysporum f. sp. lycopersici strain KACC 40038 (FOL) was generously provided by Korean Agricultural Culture Collection (KACC), Korea. Fungal and bacterial strains were grown at potato dextrose agar (PDA) and nutrient agar (NA) media, respectively.
For tomato Fusarium wilt assay, FOL was grown at PDA for 10 days at 28°C. Surface of fungal mycelia on PDA medium was filled with 5 ml of sterile distilled water (SDW), fungal spores were collected using a spreader, and filtered through cheesecloth. The spore concentration was adjusted to 1 × 106 conidia/ml by using a hemocytometer (Paul Marienfeld Superior, Lauda-Königshofen, Germany).
For potato soft rot assay, Dc was grown at nutrient broth (MBcell, Kisanbio, Seoul, Korea) medium for 2 days at 28°C. Bacterial cells were collected by centrifugation at 13,000 rpm and suspended in sterile tap water (108 cfu/ml) (OD600 of 1.0 = 109 cfu/ml).
Isolation and screening of the antagonistic activity of ANU-301
Soils were sampled from Agisan in Andong, South Korea. One gram of soil samples was suspended with 10 ml SDW, vortexed for 10 min and sieved with cheesecloth to remove large soil particle. Supernatant was serially diluted and spread on PDA medium containing 50 μg/ml streptomycin. Different fungal colonies were recovered and grown at PDA media.
Fungal isolates were screened for their antagonistic activity against different plant pathogenic fungi by using dual culture and culture filtrate assays (DCA and CFA, respectively). DCA and CFA were performed as previously described with minor modifications (Park et al., 2021; Shin et al., 2019). The pathogen and soil-isolated fungi were grown at PDA media, and agar-mycelium discs (6 mm diameter) were taken from the edge of an actively growing fungal colony for DCA. Each agar-mycelium disc was placed on PDA with 40 mm distance and incubated at 25°C for 7 days. One of the soil-isolated fungi named Andong National University-301 (ANU-301) showed significant inhibition activity against different fungal pathogens.
To prepare the culture filtrate (CF), ANU-301 was cultured in potato dextrose broth (PDB) at 25°C for 5 days, centrifuged at 13,000 rpm for 10 min, and supernatant was filtered with with 0.22 μm filters. For CFA, agar-mycelium discs of pathogens were placed in the middle of PDA media with or without 10% CF, then measured diameter of fugal mycelium after growing at 25°C for 5 days. To prepare the PDA with 10% CF, filter sterilized CF was added to autoclaved PDA after cooling to about 60°C.
For DCA and CFA, three replicates were prepared for each experiment, and plates inoculated only with the pathogen were used as control. The antagonistic activity of ANU-301 or its CF was expressed as the percentage of pathogen mycelium growth inhibition compared to the control.
Identification of ANU-301
For microscopic observation of conidiophore and conidia, ANU-301 was grown at PDA medium and observed by using the microscope (BX50, Olympus, Tokyo, Japan). The obtained image was analyzed by using ImageJ software. For molecular identification, the rDNA genes containing highly conserved internal transcribed spacer (ITS) regions were amplified by polymerase chain reaction (PCR) and sequenced. Genomic DNA (gDNA) was extracted from mycelia of ANU-301 using HiGene Genomic DNA Prep Kit (BIO-FACT, Daejeon, Korea) according to the manufacturer’s instructions. The PCR amplification and sequencing of the ITS rDNA was performed using the following primer set: ITS1: 5′-TCCGTAGGTGAACCTGCGG-3′ and ITS4: 5′-TCCTCCGCTTATTGATATGC-3′ (White et al., 1990).
PCR was performed in a 30 μl reaction mixture containing 15 μl of 2× Taq PCR Pre-Mix (Solgent, Daejeon, Korea), 1 μl of 10 pmol/μl of ITS1 and ITS4 primers, and 1 μg of gDNA. PCR amplification was carried out using thermal cycler (Multigene Gradient, Labnet, Edison, NJ, USA) by following amplification conditions: denatured at 95°C for 5 min, 33 cycles of 94°C for 30 s, 56°C for 10 s, 72°C for 40 s, and final extension at 72°C for 10 min. PCR products were analyzed by gel electrophoresis and purified using PCR purification Kit (BIOFACT) according to the manufacturer’s instructions for sequencing. The resulting sequence of the ITS rDNA of ANU-301 was analyzed with NCBI’s Gen-Bank sequence database (http://www.ncbi.nlm,nih.gov) to identify the closest species relatives and phylogenetic tree was constructed by MEGA-X software using maximum likelihood method (Kumar et al., 2018). The resulting sequence of the complete ITS rDNA of ANU-301 has been deposited in the NCBI (accession no. OL831165).
Biocontrol activity of ANU-301 in tomato and potato
Tomato (Solanum lycoperiscum L. cv. Seogwang) plants were grown in plastic tray (55 × 35 × 15 cm) containing soil mix (peat moss, perlite, and vermiculite; 5:3:2, v/v/v) at 28°C with a day length of 16 h at a light intensity of 70 μmol photons/m2/s. Six seedlings at the two-leaf stage were transplanted to a plastic pot (5 × 15 × 10 cm) containing the same soil mix. Two-week-old tomato plants were used for pathogen inoculation experiment. For biological control assay, tomato plants were uprooted and submerged into 1 × 106 conidia/ml of FOL in the absence or presence of 1 × 108 conidia/ml of ANU-301 and planted back. The infected plants were incubated in a controlled chamber at 28°C with 100% relative humidity for 2 weeks.
Potato (Solanum tuberosum L. cv. Sumi) tubers were surface sterilized by immersing sequentially in 1% NaOCl (5 min), 70% ethanol (5 min) and washed 3 times with SDW. To prepare the inoculum, 2 × 106 cfu/ml of Dc was mixed with or with our 1:50 diluted CF of ANU-301 in 1:1 ratio (Final concentrations were 106 cfu/ml Dc and 1/100 diluted CF of ANU-301). For pathogenicity assay, 10 μl of each mixture was directly injected into potato tubers by using the pipette. The inoculated potato tubers were placed in plastic container and incubated at 25°C with 100% relative humidity. The diseased area in all cases was calculated from area of the macerated region by using the ImageJ program to calculate percent disease severity. Three independent trials were carried out with three replicates. For control experiments, potato tubers were injected with 10 μl of sterilized distilled water.
Gas chromatography-tandem mass spectrometry analysis
ANU-301 was inoculation in 1 l of PDB medium for 5 days. After cultivation, broth medium was centrifuged at 10,000 ×g for 10 min and supernatant transferred to the conical flasks. In each conical flask, ethyl acetate were added to supernatant in a 1:1 ratio, shaken, and kept for overnight. Then, extraction solution were dried using a rotary evaporator. Extracts were dissolved in methanol for gas chromatography-tandem mass spectrometry (GC-MS/ MS) analysis and in DMSO for antifungal test. For GC-MS/MS analysis, 5977A Series GC/MSD System (Agilent Technologies, Santa Clara, CA, USA) was used. Temperature program of 40°C (1 min hold) to 300°C (10 min hold) at a rate of 10°C/min. The injector and mass interface temperature was 300°C, and the gas flow rate of the carrier gas, helium, was 1.0 ml/min. Injection mode was split and mass range was 50 to 400 (m/z). The result was analyzed using the National Institute of Standards and Technology spectral library version 11 (NIST 11 spectral library).
Results
Screening of fungi with antifungal activity
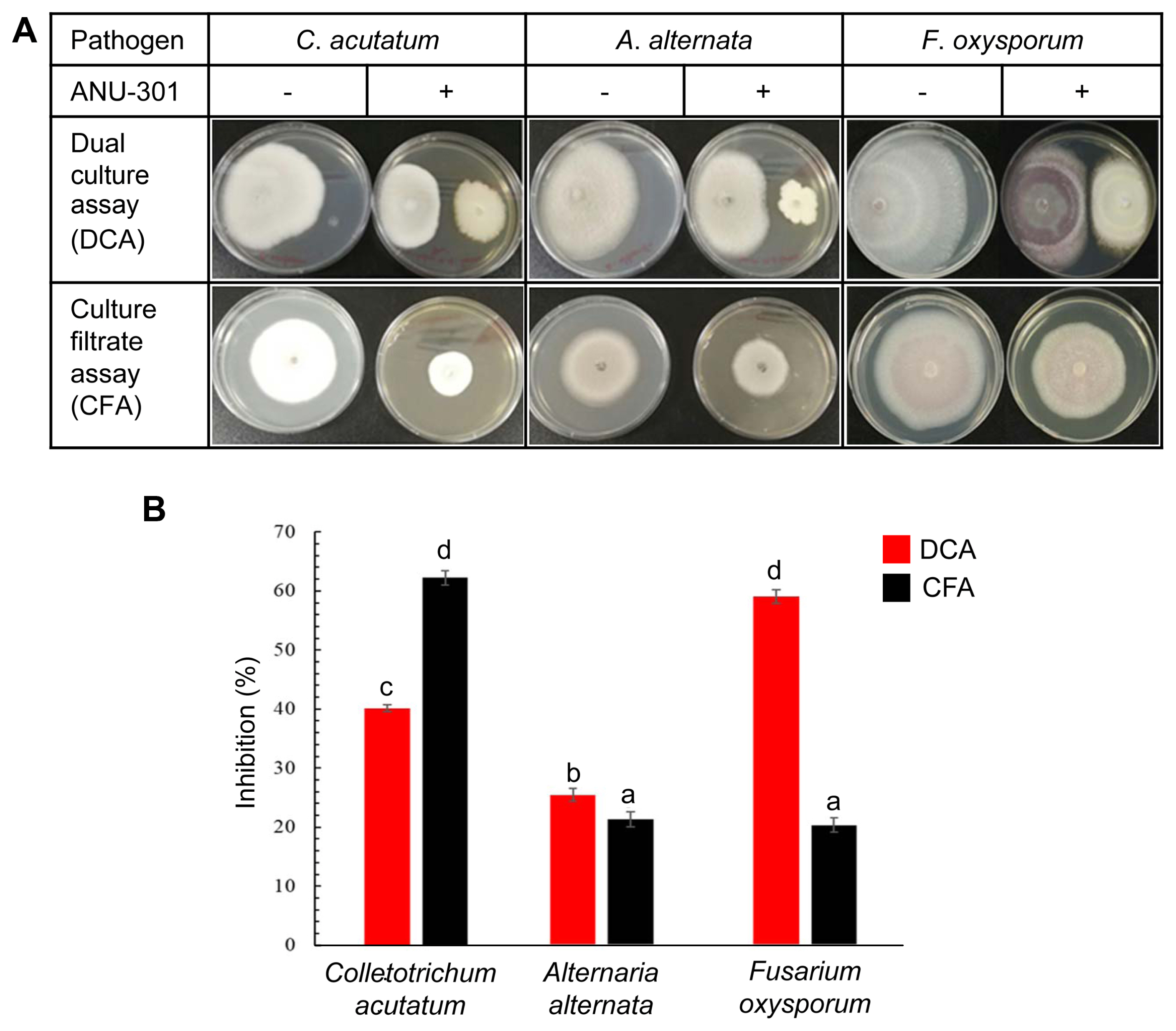
To screen the soil fungi with biocontrol activity against different plant pathogens, unknown fungal isolates were recovered from the soil by plating on PDA media, then screened by DCA and CFA (Fig. 1). One of the fungal isolate, ANU-301, showed distinct growth inhibition activity (GIA) against different plant pathogenic fungi, such as C. acutatum, A. alternata, and F. oxysporum, in DCA and CFA (Fig. 1A). In DCA, GIA of ANU-301 against F. oxysporum was about 58.5%, then followed by C. acutatum (40.1%) and A. alternata (27.3%) (Fig. 1B). By contrast, ANU-301 showed the highest GIA against C. acutatum (62.5%), whereas much lower GIA against A. alternata (21.7%) and F. oxysporum (20.2%), in CFA. Although ANU-301 showed different levels of antifungal activity against different plant pathogens, it seems likely have broad spectrum control efficiency against different fungal pathogens.
Identification of ANU-301
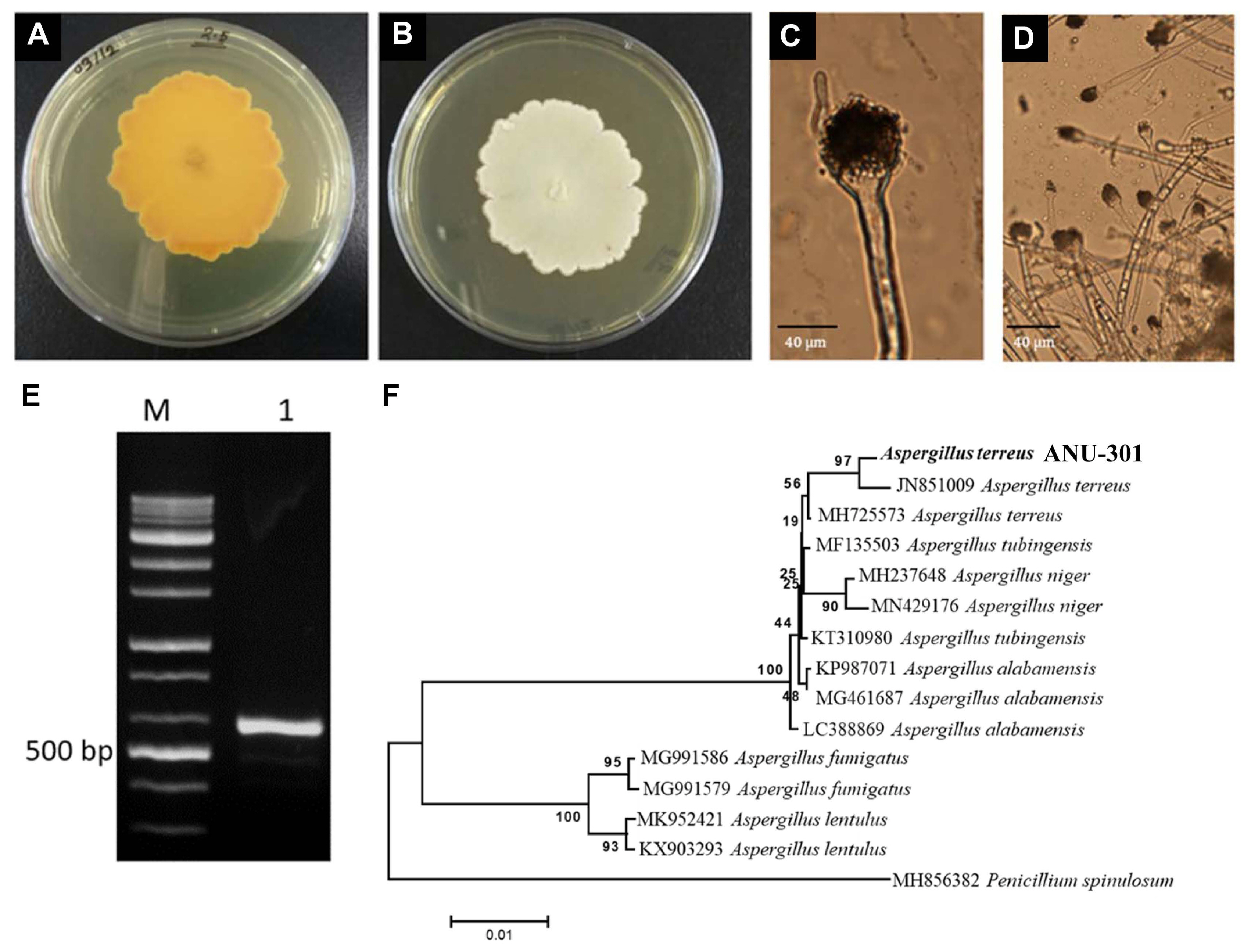
To identify the ANU-301, morphological characteristics under the microscope and ITS sequencing-based phylogenetic analysis was performed (Fig. 2). ANU-301 incubated in PDA grew well and produced white-colored mycelial mat (Fig. 2A and B). Under the microscope, mycelia have a septum and asexual conidia were observed arising from globose-shaped conidiophores, which is a characteristic of Aspergillus spp. (Fig. 2C and D). To further identify the ANU-301, ITS region was amplified by PCR and sequenced for phylogenetic analysis (Fig. 2E and F). From the analysis, ANU-301 showed the highest sequence homology with A. terreus and formed distinct clade from the other Aspergillus spp. Based on these procedures, it is named as A. terreus isolate ANU-301 (ANU-301 from now on).
Biocontrol activity of ANU-301 against tomato Fusarium wilt disease
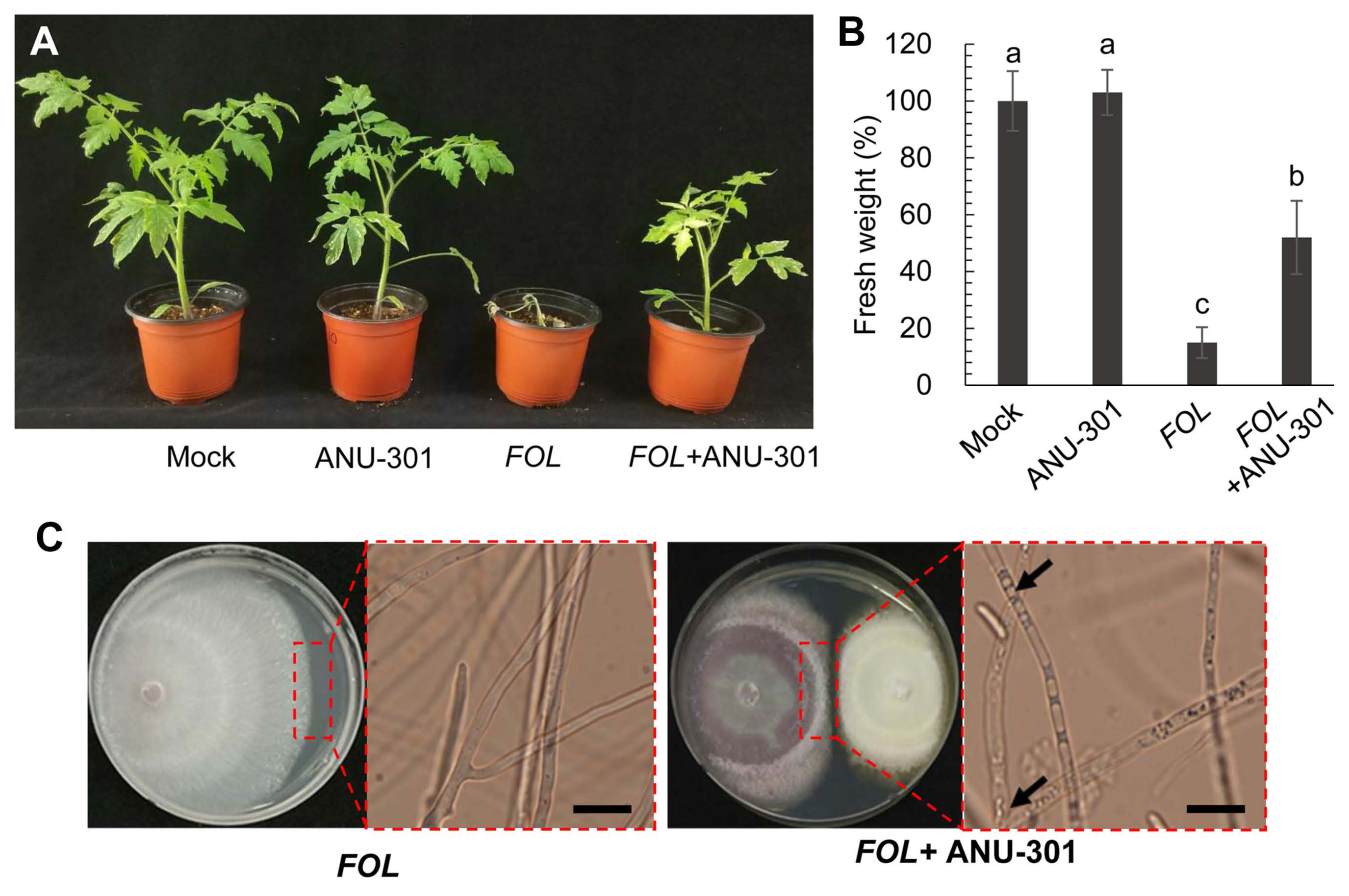
As ANU-301 showed the highest GIA against FOL in DCA, its biocontrol activity against tomato Fusarium wilt disease was examined (Fig. 3). Two-week-old tomato plants were inoculated by submerging the roots in 106 conidia/ml of FOL with or without 108 conidia/ml of ANU-301. Mock or ANU-301 treated plants showed no significant differences in growth rate; however, tomoto plants showed very severe growth retardation and wilting phenotype at 2 weeks after inoculation with FOL (Fig. 3A and B). Importantly, tomato plants co-inoculated with FOL and ANU-301 showed significantly reduced disease symptoms.
To confirm whether ANU-301 has direct effect, mycelial growth of FOL in the absence or presence of ANU-301 was observed microscopically (Fig. 3C). Abnormal large vesicle formation was observed inside the hyphae of FOL at the contact point with ANU-301, and purple-colored pigmentation of FOL mycelium was observed during the DCA. These findings suggest that ANU-301 somehow alters physiology of FOL, thereby reducing their growth. Taken together, ANU-301 not only showed antifungal activity against FOL in vitro, but also successfully suppressed tomato Fusarium wilt disease in pot experiments.
Biocontrol activity of ANU-301 against potato soft rot disease
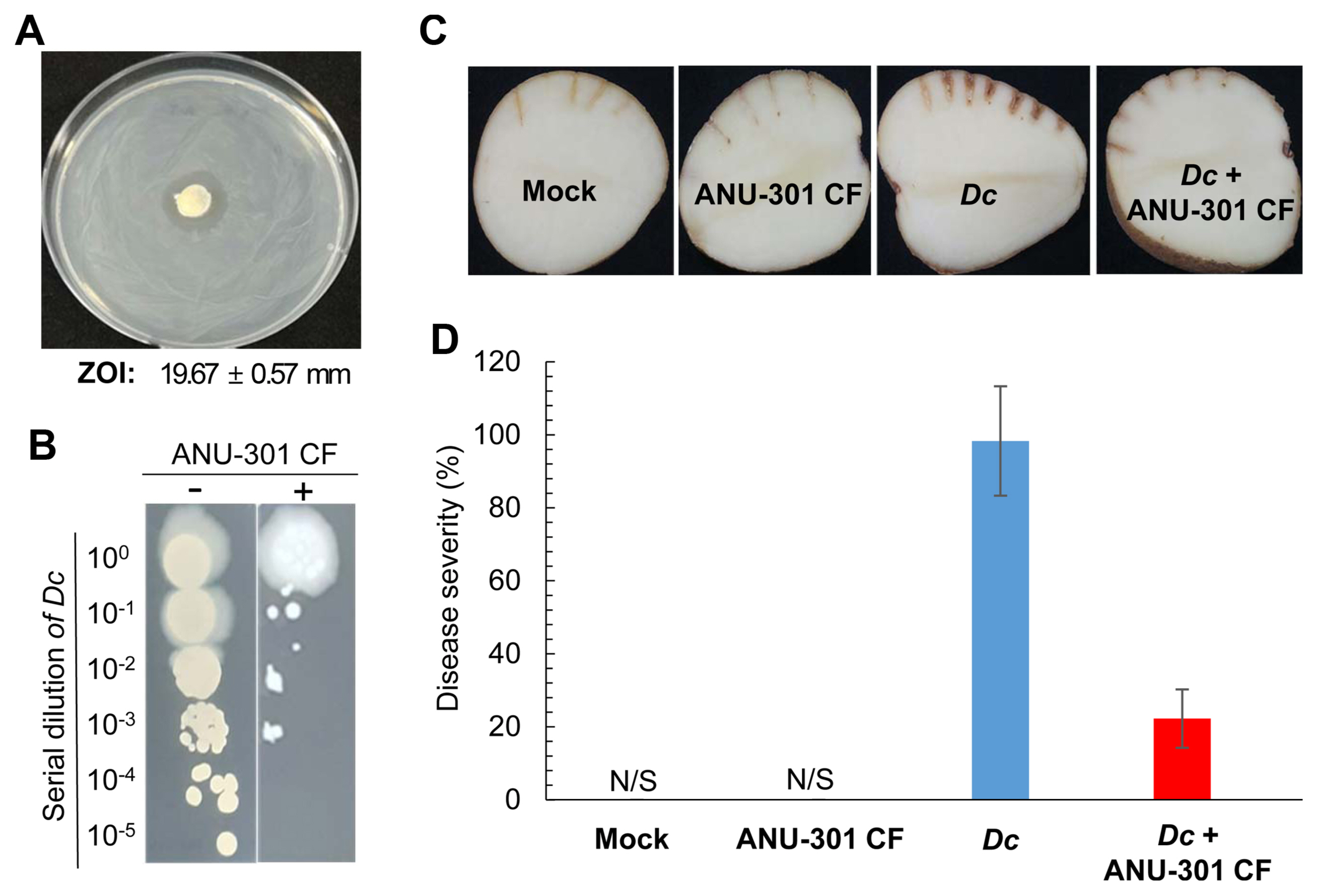
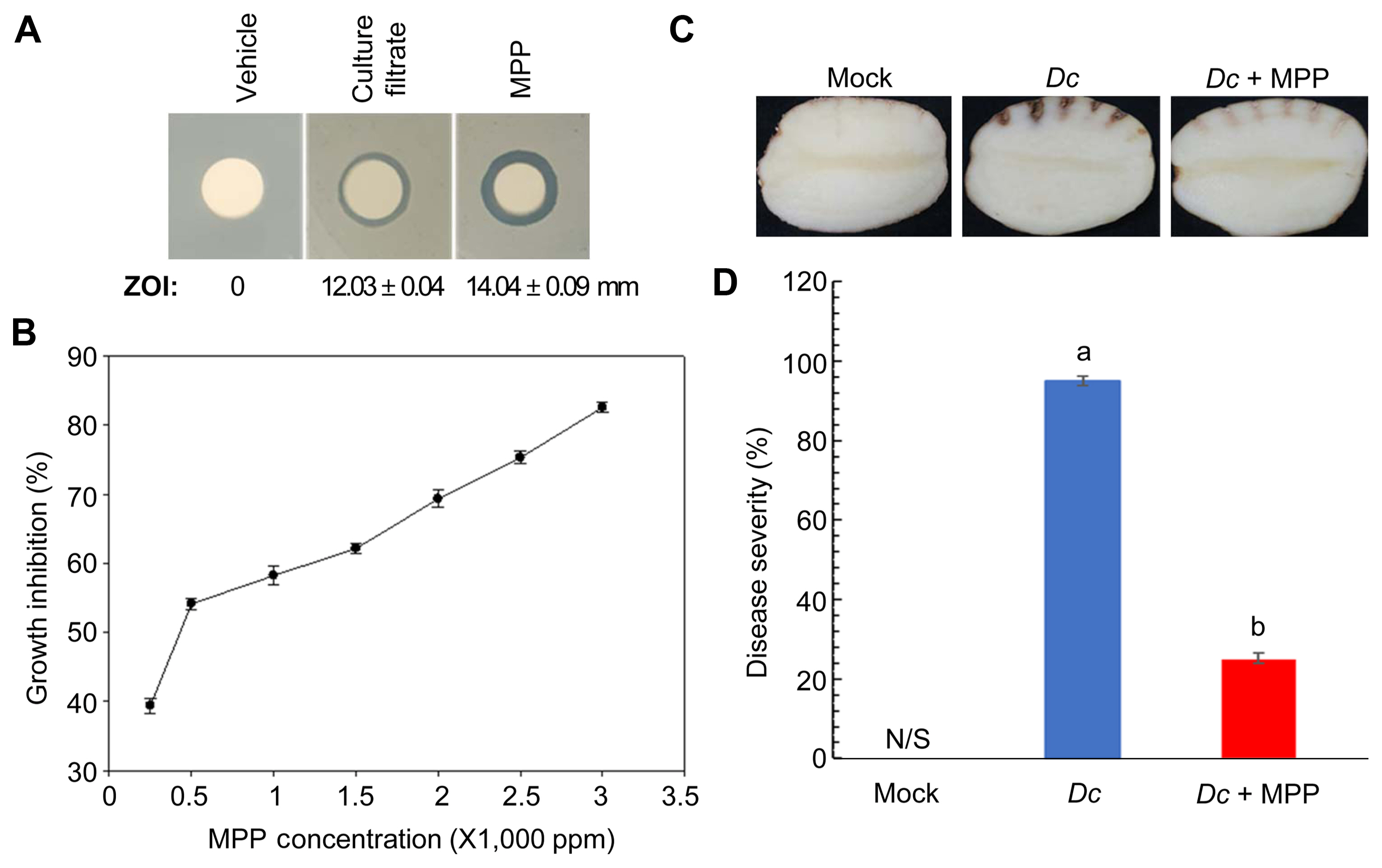
To further examine whether ANU-301 has antibacterial activity, agar plug diffusion assay was performed against Dc, a causal agent of potato soft rot disease. Interestingly, ANU-301 also showed antibacterial activity as developed clear zone in Dc grown NA media (Fig. 4A). Diameter of zone of inhibition (ZOI) was 19.67 ± 0.57 mm. Antibacterial activity of CF of ANU-301 was examined. In this assay, 1:100 diluted CF of ANU-301 was mixed with 106 cfu/ml of Dc, incubated for 1 h, then serially diluted and plated on NA media. As shown in Fig. 4B, CF of ANU-301 successfully suppressed the growth of Dc, suggesting ANU-301 also has antibacterial activity against Dc.
As CF of ANU-301 showed antibacterial activity against Dc, its biological control activity against potato soft rot disease was also analyzed. As expected co-treatment of CF of ANU-301 significantly reduced disease symptom developed by Dc in potato tuber (Fig. 4C and D). In mockand ANU-301 CF-treatd potato, none of visible symptoms were observed, whereas Dc-inoculated potato developed soft rot symptoms. Co-treatment with ANU-301 CF significantly reduced the diseased severity in potato by 79.1% (Fig. 4D). Taken together, ANU-301 showed biological control activity against two distinct plant pathogens, FOL and Dc.
Identification of MPP as a major antibacterial secondary metabolite in the CF of ANU-301
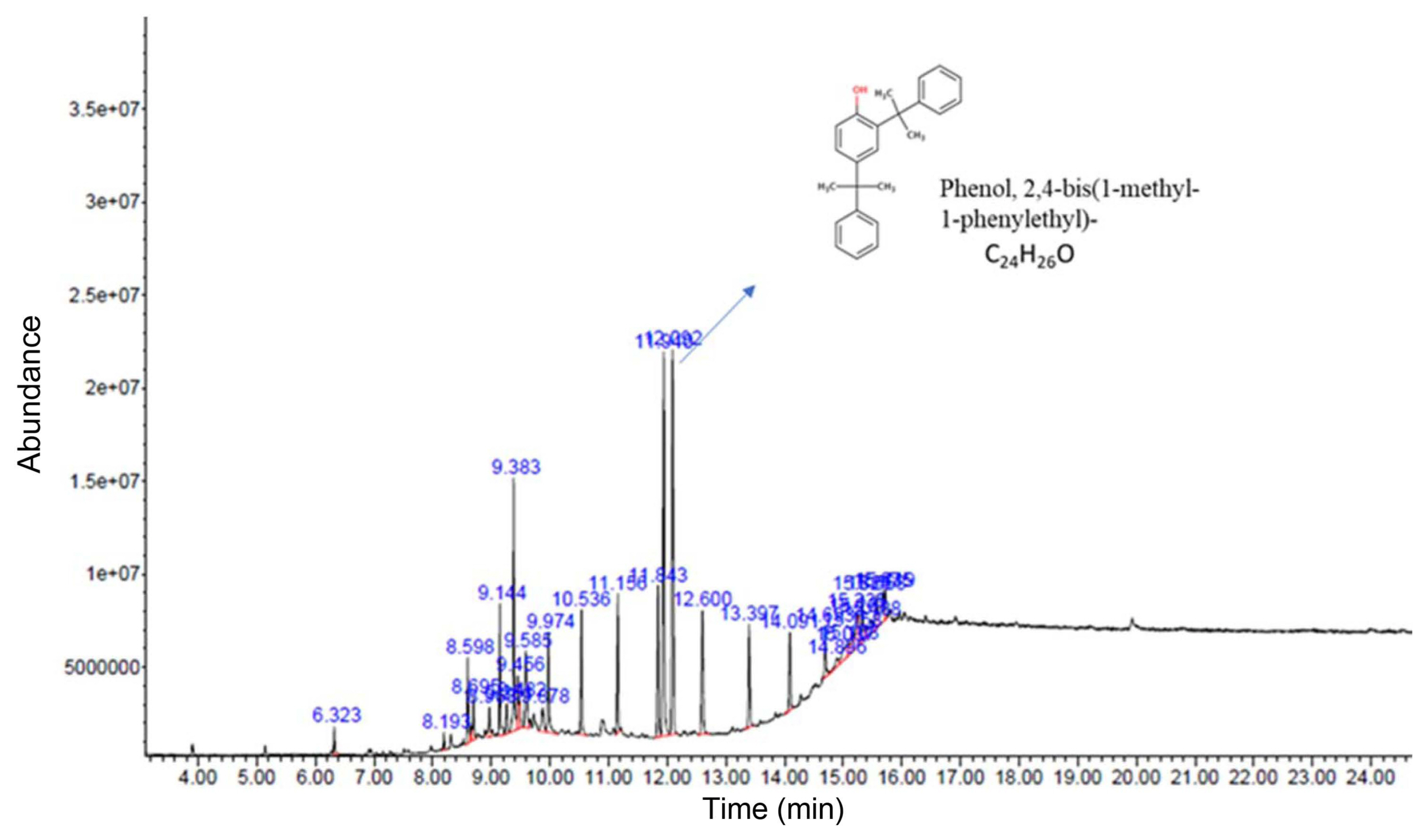
To further examine the major antimicrobial secondary metabolites in CF of ANU-301, GC-MS/MS experiment was performed. From the different compounds identified, MPP was the most abundant from the CF of ANU-301 (abundance: 15.05%), and followed by 2,3,4,5-tetraphenyl-1H-pyrrole (abundance: 14.01%), 2,6-bis(1,1-dimethylethyl)-4-(1-methyl-1-phenylethyl)-phenole, octadecane and tatracosane (Table 1, Fig. 5).
MPP successfully suppressed bacterial growth of Dc, in NA media as developed clear zone in filter disk diffusion assay (10 μl of 400 ppm MPP or 1/100 diluted CF were used) (Fig. 6A), but not of FOL (Supplementary Fig. 1). MPP-induced diameter of ZOI was 14.04 ± 0.09 mm. Bacterial growth was measured by using the spectrophotometer at OD600 in the presence of different concentrations of MPP (Fig. 6B). MPP inhibited growth of Dc in a concentration-dependent manner with a calculated IC50 of 505.9 ppm (~1.5 mM). Biocontrol activity of MPP against potato soft rot disease was examined. As expected, co-inoculation of Dc with MPP significantly reduced disease severity in potato tuber (Fig. 6C and D). Mock-inoculated potato developed negligible browning at the wounded site, whereas Dc-inoculated potato developed severe browning and soft rot symptoms (disease severity of 89.8%). Co-treatment of MPP significantly reduced disease severity by 22.1%. Taken together, major secondary metabolite MPP of ANU-301 CF was identified to have antibacterial activity.
Discussion
Aspergillus, a well-known saprophytic filamentous fungal genus, play different biological roles in different niche, with Aspergillus niger and Aspergillus flavus being the most studied and significant species for their secondary metabolites (Hossain et al., 2017; Park et al., 2017; Yu et al., 2021). A. terreus, a less well-known and explored microbe, was recently discovered to be a significant biological control agent and plant growth promoting microorganism (Frisvad and Larsen, 2015; Vassileva et al., 2020). The biological control capabilities of A. terreus (ANU-301) against several plant diseases are addressed in this work.
The closely related phenolic compounds, such as MPP, 2,4-Di-tert-butylphenol (2,4-DTBP), and 2,6-bis(1,1-dimethylethyl)-4-(1-methyl-1-phenylethyl)-phenol, are known to be toxic secondary metabolite produced by various group or organisms (Melappa et al., 2017; Shaaban et al., 2021; Zhao et al., 2020). 2,4-DTBP is a lipophilic phenol that has been identified as a lethal secondary metabolite in 169 species of organisms, including A. terreus (Aissaoui et al., 2019; Akshatha et al., 2016; Belghit et al., 2016; Lenartowicz et al., 2015; Varsha et al., 2015; Viszwapriya et al., 2016; Zhang et al., 2008; Zhao et al., 2020). 2,4-DTBP is efficient against the agriculturally significant rootrot fungus F. oxysporum by suppressing spore germination and hyphal outgrowth through a possible interaction with β-tubulin in microtubules (Dharni et al., 2014). 2,4-DTBP also known to inhibit the production of quorum sensing (QS)-mediated biofilm formation, while it increases the hydration of the cell wall, resulting in decreased biofilm formation (Padmavathi et al., 2014). Intercellular communication in bacteria, technically referred to as QS, is a critical factor in disease spread and pathogenesis because it regulates biofilm formation. Reportedly, it plays an important role in development of Dickeya-induced Potato soft rot (Baltenneck et al., 2021). ANU-301 was also identified as a source of MPP and 2,6-bis(1-methyl-1-phenylethyl)-phenol by GC-MS/MS analysis as the first and third most abundant chemical compounds, respectively. Unexpectedly, MPP did not show significant antifungal activity against FOL (Supplementary Fig. 1). In accordance with this result, CF of ANU-301 showed much lower antifungal activity against FOL in CFA compared to its antifungal activity in DCA (Fig. 1). This may suggest that (1) antifungal activity of ANU-301 against FOL is not dependent on MPP production, and/or (2) ANU-301 can produce specific antifungal compounds against FOL in the presence of the pathogen (Note that CF of ANU-301 prepared from the pathogen-free media show little antifungal activity against FOL, but still effective to restrict the growth of FOL in DCA).
In GC-MS/MS analysis, we discovered 2,3,4,5-tetraphenyl-1H-pyrrole as the second highest peak from the analysis of CF of ANU-301. Pyrrole is a five-membered aromatic ring having the chemical formula C4H5N (Ahmad et al., 2018). Pyrrole is a fundamental building block of biochemistry due to its flat electron-rich ring, which is particularly vulnerable to electrophilic assault and may react with a wide variety of biomolecules through hydrogen bonding and π-π stacking interactions (Domagala et al., 2015). Analogs of pyrrole are being investigated as possible sources of antifungal and antibacterial activity (Bhardwaj et al., 2015). The antibacterial activity of this compound is controlled by substituting alternative molecules, scaffolds, or moiety, groups in the pyrrole ring following attached analogs, including the phenyl (diaryl) moiety (Wang et al., 2011; Arumugam et al., 2012; Bhosale et al., 2013; Jana et al., 2005; Raimondi et al., 2006; Thach et al., 2016; Varshney et al., 2017). 2,3,4,5-tetraphenyl-1H-pyrrole is a pyrrole derivative with four phenyl rings that has recently been shown to have antibacterial (mycobacterial) activity (Bhardwaj et al., 2015).
As fourth and fifth peak in GC-MS/MS analysis ANU-301 shows octadecane and tetracosane respectively. Octadecane (C18H38) and tetracosane (C24H50) are aliphatic hydrocarbon (alkane) maintaining its general formula CnH2n+2, is a dangerous environmental toxic chemicals in petroleum-contaminated soil consisting of recalcitrant compound make it non-degradable (Bajagain et al., 2018; Campo et al., 2017; Hamouda et al., 2016; Hussain et al., 2018; Meckenstock et al., 2016). Some recent reports proved that they also have significant antimicrobial properties (Abiona et al., 2019; Barupal et al., 2019; Bouhlal et al., 2020; Li et al., 2021; Rajaofera et al., 2019; Ramírez et al., 2021; Wang et al., 2020).
Tomato is a widely grown vegetable crop in the Solanaceae family with high nutritional content and antioxidant effects (Nahar and Ullah et al., 2012). Fusarium wilt is strongly related with tomato production loss in field conditions (Chakraborty and Acharya, 2017). Blackleg and soft rot in potato, both caused by D. chrysanthemi, are among the most damaging bacterial diseases plaguing this world’s fourth most important food crop (Pédron et al., 2021). The potentials of Aspergillus spp., particularly A. terreus, are currently underutilized (Galeano et al., 2021; Vassileva et al., 2020). Our investigation established its potential as a future contender for effective biological control agent.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement
Supplement Print
Print






