



Mulberry (Morus spp.) is a perennial fast growing woody plant, which is widely cultivated in many tropical and subtropical areas. The leaves of the mulberry are economically important as they are the only source of feed for the monophagous silkworm (Bombyx mori L.). After China, India is one of the leading producers of mulberry with a total acreage of around 239,926 hectares. Karnataka, Andhra Pradesh, West Bengal and Tamil Nadu are the major silk producing states in India (Central Silk Board, 2020). The tropical climate of South India is highly favorable for mulberry cultivation where the crop is harvested at every 70 days (Gnanesh et al., 2021). Mulberry production is limited by several diseases as the perennial nature of the mulberry provides scope for long-term survival and multiplication of soil-borne pathogens (Sharma et al., 2003). Among the soil-borne diseases, root rot is epidemic in nature and a major limiting factor in the cultivation of mulberry. Root rot causes 30% mortality of plants with a 14% decrease in leaf yield, besides deteriorating the leaf quality (Chowdary, 2006; Rajeswari and Angappan, 2018).
In mulberry, different kinds of root rot have been reported, such as black root rot, dry root rot, charcoal root rot, violet root rot, white root rot, Armillaria root rot, Rhizopus rot, and bacterial root rot (Gnanesh et al., 2021; Radhakrishnan et al., 1995; Sharma et al., 2003; Yoshida et al., 2001). Amongst them, black root rot (BRR) caused by Lasiodiplodia theobromae (syn. Botryodiplodia theobromae), dry root rot (Fusarium solani and F. oxysporum), and charcoal root rot (Macrophomina phaseolina) are frequently reported in India (Pinto et al., 2018).
Lasiodiplodia species are harmful fungal pathogens and have been associated with nearly 500 plant hosts causing various degrees of crop losses. It causes root rot disease in mulberry that affects all parts of the plant and it spreads rapidly affecting a large number of plants in a short period leading to the abandonment of mulberry gardens (Pappachan et al., 2020). The fungus prefers 25-30°C for good growth and survives on all types of substrates. After pruning, the pathogens enter the host through the cut ends of the stem. Once, the plant becomes vulnerable to infection, the fungus dominates inside the roots by multiplying the hyphae rapidly in the cortical tissues and extending up to the pith region. It enters the xylem vessels and causes the death of the plants (Sharma et al., 2003). Diseased plants appeared stunted with dark colored stems and chlorotic leaves, gradually the leaves get wilted and defoliated prematurely. The fine feeder roots and the main root system of these infected plants will damage severely with typical symptoms like discoloration of root xylem followed by the decay of root cortex. Also, the bark of the roots becomes fragile and smelled emanating a foul odour. Gradually as the damaged roots could not firmly hold the plant in the soil they can be easily uprooted (Sowmya et al., 2018; Xie et al., 2014).
For the first time in China, BRR caused by L. theobromae was reported by Xie et al. (2014) and it has a serious impact on the sustainable growth of the local sericulture industry (Xie et al., 2016). Similarly, Sowmya et al. (2018) studied the severity of the BRR disease in major mulberry growing areas of India. The Lasiodiplodia species is known to cause dieback, cankers, and stem and root rot in multiple crops like almonds, blueberry, castor, Chinese hackberry, citrus, cocoa, coconut, grapevines, groundnut, jackfruit, mango, melon, olive, Pinus spp., strawberry, and rice (Bautista-Cruz et al., 2019; Chen et al., 2021; Liang et al., 2019; Nam et al., 2016; Pečenka et al., 2021; Rosado et al., 2016; Saeed et al., 2017).
As it is not sufficient to classify Lasiodiplodia species based on morphological features, DNA sequence-based approaches have been widely recommended (Bautista-Cruz et al., 2019). For many years, L. theobromae was treated as a monotypic genus within the Botryosphaeriaceae (Slippers et al., 2013). However, phylogenetic analysis evidenced the existence of many additional species (de Silva et al., 2019).
Various chemical and biological methods were recommended to manage the disease caused by Lasiodiplodia spp. and avoid economic losses (Kamil et al., 2018). The chemical control measures are non-judicious and undesirable due to the buildup of resistance among pathogens (Leroch et al., 2011). They are also unsafe for the environment and human health and toxic to silkworms (Naik et al., 2010). Biocontrol measures were not very effective due to the influence of various factors on their efficacy (Nelson, 2004). Genetic improvement by breeding a resistant variety ensures a cost-effective and environment-friendly system for the control of plant diseases (Arunakumar et al., 2021; Pandey et al., 2021). Identification and breeding of resistant mulberry varieties is the only justifiable option to achieve sustainable mulberry cultivation.
There are limited data on BRR of mulberry in India, and the previous characterization of L. theobromae was based on morphology. Sowmya et al. (2018) used random amplified polymorphic DNA (RAPD) and simple sequence repeats (SSRs) to study the genetic variability, among the ten isolates of L. theobromae causing BRR of mulberry, similarly, Pappachan et al. (2020) characterized only one isolate of L. theobromae. However, these studies could not reveal the molecular phylogeny of isolates and did not attempt to identify resistant sources to L. theobromae. Several research investigations reported new species of Lasiodiplodia (de Silva et al., 2019; Rosado et al., 2016) indicating the extension of its host range highlights the need for prospecting novel Lasiodiplodia species associated with mulberry. Accurate detection of pathogens is very essential for the development of proper management approaches, moreover use of highly pathogenic or aggressive isolates is necessary for inoculation trials for selecting germplasm accessions with broader resistance to diseases (Oliveira et al., 2021). Also, there is an immediate need to identify resistant sources to transfer resistance genes into elite backgrounds of mulberry.
Thus, the objectives of the present study were as follows: (1) to characterize L. theobromae causing BRR of mulberry in India, morphologically, combined with DNA sequence analysis using internal transcribed spacer (ITS) region and β-tubulin, (2) to evaluate the pathogenicity of the selected isolates (3) finally, to identify BRR resistant resources and their utilization in future mulberry breeding programs to develop root rot-resistant varieties.
Materials and Methods
Sample collection and isolation of the fungus
Seventy four diseased root samples of mulberry were collected from farmers field from 2017 to 2019 in high-incidence states (Andhra Pradesh, Karnataka, Tamil Nadu, and Telangana) of South India. The root rot causative fungal pathogen was isolated following the root bit method of isolation. The infected mulberry roots showing typical root rot symptoms were chopped into small bits and surface-sterilized using 5% sodium hypochlorite for 2 min. These root bits were washed with sterile water and dried up on a filter paper (Whatmann No. 1). Dried root bits were placed on Petri plates with sterile potato dextrose agar (PDA; HiMedia, Mumbai, India) under aseptic conditions. The plates with root bits were incubated at 27 ± 1°C for 7 days to obtain fungal growth. Pure cultures were obtained by sub-culturing matured hyphae on 2% water agar plates and stored for further use at 4°C.
Morphological identification of fungal isolates
The morphology of 7-day-old fungal cultures grown on PDA medium incubated at 27°C under dark conditions was examined. Cultural characteristics like colony color, pigmentation, texture, and growth were noted. Conidial morphology was observed using light microscopy and conidial dimensions of 20 selected isolates were determined (Supplementary Table 1). The length and width of 10 mature and immature conidia per isolate were measured using ImageJ software.
Scanning electron microscopy
Glutaraldehyde (2.5%) prepared in 0.2 M cacodylate buffer (pH 7.2) was used for the initial fixation of fungal cultures. The fixed cultures were washed using cacodylate buffer and then by double distilled water dehydrated in ethanol series. Critical drying of the dehydrated samples was carried out by using a Critical Point Dryer (EMS850, Electron Microscopy Sciences, Hatfield, PA, USA). Lastly, samples were coated with gold nanoparticles in Sputter Coater (EMS550, Electron Microscopy Sciences) mounted onto copper stubs using double side sticky tape. Microscopic images were obtained after scanning under JEOL 100 CX ASID-4D scanning electron microscope (JEOL Ltd., Tokyo, Japan) at 20 kV (Arunakumar et al., 2018; Bozzola and Russell, 1992).
Molecular identification of the pathogen DNA isolation and polymerase chain reaction amplification
Pure cultures of 20 fungal isolates were inoculated in 50 ml of sterile potato dextrose broth and incubated for 7 days at 27 ± 1°C, without shaking. The mycelial mats were separated by using sterilized filter paper (Whatmann No.1). DNA extraction was performed, following the method described by Gnanesh et al. (2021).
The representative fungal isolates of mulberry were identified by sequencing ITS region, ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATAT GC-3′) primers (White et al., 1990) and β-tubulin region, βt2a (5′-GGTAACCAAATCGGTGCTG CTT TC-3′) and βt2b (5′-ACCCTCAGTGTAGTGACCCTTGGC-3′) (Glass and Donaldson, 1995). The polymerase chain reaction (PCR) amplification was performed with a thermocycler (GeneAmp PCR system 9700, Applied Biosystems, Foster City, CA, USA). in a 25 μl reaction mixture consisting of 2 μl of template DNA (40 ng), 12 μl of 2× Ampliqon master mix, 1.5 μl of each primer (10 pM) synthesized from Eurofins, Bangalore, India and 8 μl of nuclease free water. The PCR parameters for ITS gene PCR amplification: initial denaturation at 94°C for 4min followed by 35 amplification cycles of 94°C for 30 s, 55°C for 30 s and 72°C for 1 min, and final extension at 72°C for 8 min. Similarly, β-tubulin gene PCR amplification was performed in a 25 μl reaction mixture which contained 10 μl of 2× Ampliqon master mix, 2 μl template DNA (40 ng), 11 μl of nuclease free water and 1 μl of each primer (10 pM). The PCR parameters for partial β-tubulin region are initial denaturation at 95°C for 5min, followed by 35 amplification cycles of 94°C for 45 s, 55℃ for 45 s, and 72℃ for 1 min, and final extension at 72°C for 7 min. Amplified products were electrophoresed using 1.5% agarose gel stained with ethidium bromide with 1× TAE running buffer, and visualized in a UV transilluminator. A 100 bp ladder was used as a marker and gels were documented using GelDoc Bio Imaging System (Syngene, Cambridge, UK). The amplified PCR products were purified and sequenced by Sanger sequencing method, Eurofins Pvt., Ltd. (Bengaluru, India).
Phylogenetic analysis
Sequence alignments of ITS and β-tubulin were carried out using the MEGA X software (Kumar et al., 2018). The obtained nucleotide sequences were edited using BioEdit software and submitted to BLAST search analysis to identify the sequence homology in NCBI database. Finally, the sequences were deposited in GenBank and obtained the accession numbers. The phylogenetic analysis of concatenated data was done by selecting the best nucleotide substitution model using Bayesian Information Criterion (BIC) in Mega X software (Kumar et al., 2018). Tamura 3-parameter (T92) model was selected for phylogeny construction and the tree was rooted to Barriopsis fusca as an outgroup. Type strain and reference sequences were obtained from GenBank and the trees were analyzed by the bootstrap method with 1,000 replications.
Pathogenicity test
Pathogenicity tests of 20 representative isolates were performed on two susceptible mulberry accessions, Victory-1 and Thailand Male, following the root dip method under glasshouse conditions (Arunakumar and Gnanesh, unpublished). Four months old saplings grown in raised nursery beds were gently uprooted and dipped in the conidial suspension (1 × 106 spore load per ml load) for 12 h. Control plants were also dipped in sterile distilled water for 12 h. Later, saplings were transplanted into the earthen pots having sterile soil arranged with three replications following a completely randomized design (CRD). The infected plants were watered regularly to maintain sufficient moisture. BRR symptoms, wilting, and rotting percentage were recorded after 120 days of post inoculation. The pathogenicity of the isolates was classified using the wilting and rotting percentage as described in Table 1.
Screening of diverse mulberry germplasm accessions
Forty five mulberry germplasm accessions including exotic and indigenous collections were used for screening. The accessions represent eight different countries including India (33), Indonesia (3), Japan and Thailand (2 accessions each), Philippines, Australia, China and France one accessions each. Among the 45 accessions 17 accessions belonged to collections, eight were open pollinated hybrids, six were from the survey, five accessions were from selections, and four each accession were from cross pollinated hybrids and clonal selection respectively and only one accession was developed. These accessions belonged to six different Morus spp. including a known popular susceptible variety Victory-1, were chosen for identification of resistance against L. theobromae causing BRR of mulberry (Table 2). Four months old saplings of each accession were uprooted gently and inoculated following the root dip method as mentioned above. The inoculated pots with four replications (single plant/replication) were arranged in a CRD. The experiment was conducted under glasshouse conditions with two control plants free from inoculation and repeated twice. The aerial observation of the total number of leaves, shoots (wilted and healthy), and the number of plants dead was recorded at 30 days intervals in each accession. Plants were uprooted after 120 days of post inoculation and observations on weight of the whole root, healthy root and above ground biomass per plant (g) were recorded. These disease indices like wilting and rotting percentages were calculated according to formulae described by Sowmya et al. (2018). The disease severity index and disease reaction were categorized for each accession based on the wilting and rotting percentage using the modified scale as mentioned in Table 1 (Ahmed et al., 1999).
Statistical analysis
The collected data were statistically processed using IBM SPSS Statistics software version 23.0 (IBM Corp., Armonk, NY, USA). One-way ANOVA analysis was performed to study the difference in the pathogenicity of the selected L. theobromae isolates and means of the BRR disease scores were compared using Tukey test (P < 0.05).
Results
Fungal isolation and morphological identification
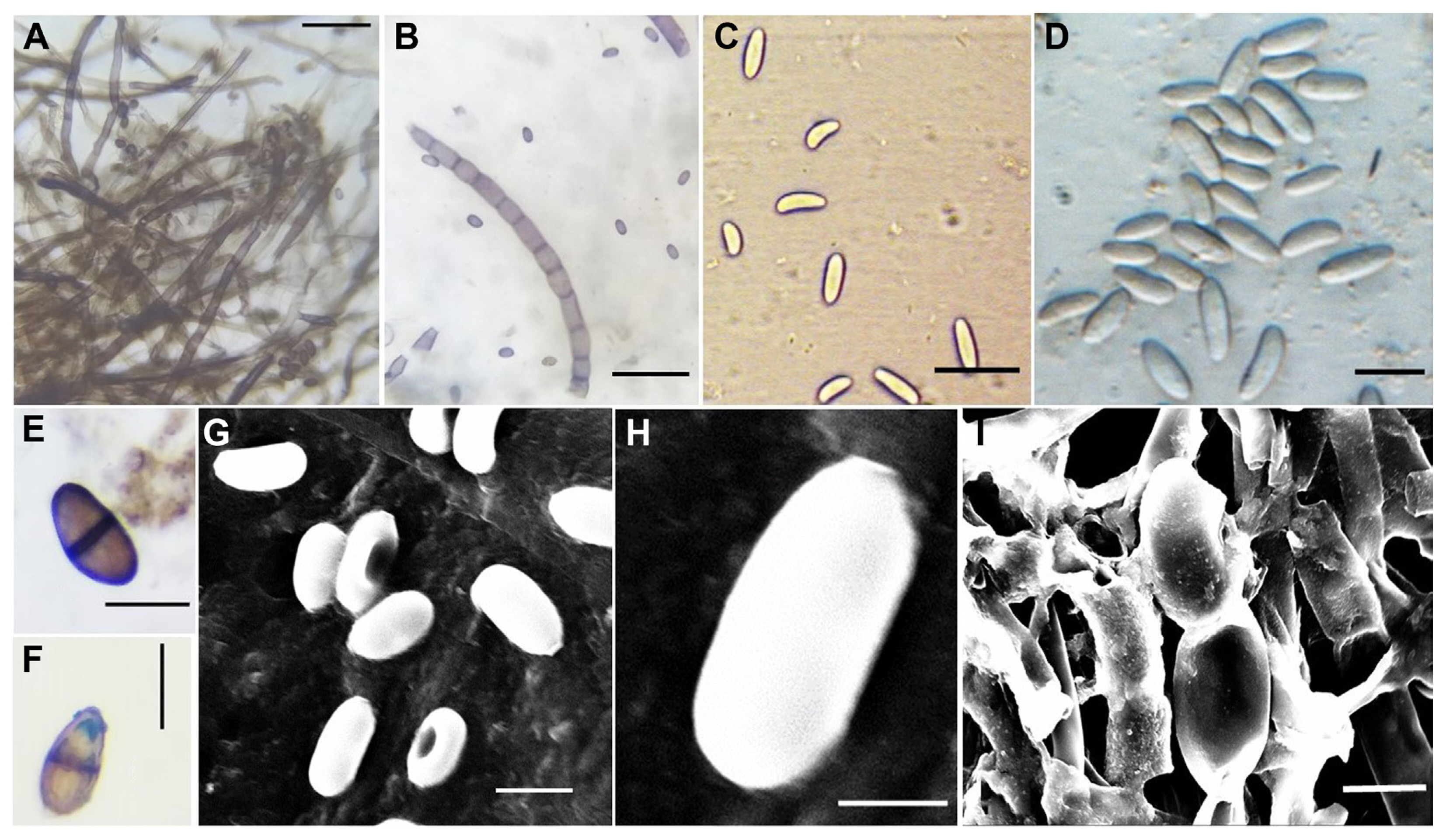
Symptomatic mulberry plants in the field exhibited varying degrees of BRR symptoms, characterized by yellowing and wilting of leaves (Fig. 1A), browning of the severely infected roots and in some cases appearance of newly regenerated roots were observed (Fig. 1B). The stem of the symptomatic plants was browning of root xylem and rotting of root cortex (Fig. 1C). Fifty seven isolates of L. theobromae were successfully recovered from 74 root rot samples collected from Southern India. The isolated fungi were grown on PDA media at 27°C in dark conditions. The cultures showed initially off-white to grey with fluffy, round to irregular margin, flat aerial mycelia were grown radially and uniformly in all directions and covered the surface of the media (90 mm diameter Petri dishes) within 4-5 days after incubation (Fig. 2A). Initially, the reverse side of the culture showed algae green dot patches and longitudinal striations towards the periphery. After 14-15 days of incubation, the reverse side turned to black color (Fig. 2B) and the upper surface formed dark grey colonies with fluffy aerial mycelium (Fig. 2C). Liquid exudates were produced and became dry after a few days (Fig. 2D).
The virulent isolate MRR-142 is shown in the microscopic pictures in Fig. 3. The mycelia were hyaline and septate, and some of the malformed mycelia were also observed (Fig. 3A and B). Pycnidia were semi-immersed, solitary, globose, papillate, and leaden-black forms after 15-20 days of incubation. Paraphyses were cylindrical, aseptate and hyaline. Conidiogenous cells were characterized as holoblastic, cylindrical and hyaline. Conidia produced after 15-20 days of incubation, immature conidial length measured from 13.34 μm to 7.4 (± 1.24) μm and breadth were ranged from 4.47 μm to 2.28 (± 0.51) μm (Fig. 3C and D). The mature conidial length was found in the range from 25 μm to 17 (± 3.0) μm and breadth was in the range from 14 μm to 7 (± 2.0) μm (Supplementary Table 1). Conidia were initially hyaline, aseptate, ellipsoid to obovoid, unicellular, cinnamon to sepia, dense walled and rounded with longitudinal light striations, then mature conidia form 1-2 septa (Fig. 3E and F). The conidial morphology and size indicated that the isolates were L. theobromae. The fine structure of conidial surfaces and conidia on a conidiogenous cell was observed using scanning electron microscopy (SEM). Conidial surface morphology of L. theobromae, although appeared to be smooth when viewed in a light microscope they are roughened and verruciform when viewed in high magnification in SEM (Fig. 3G-I). The ornamentation type could not be reliably distinguished with light microscopy. Conidia are broadly ovoid to ellipsoidal, presenting both ends broadly rounded or end rounded and the base narrowed or apiculate (Fig. 3H).
Phylogenetic analysis
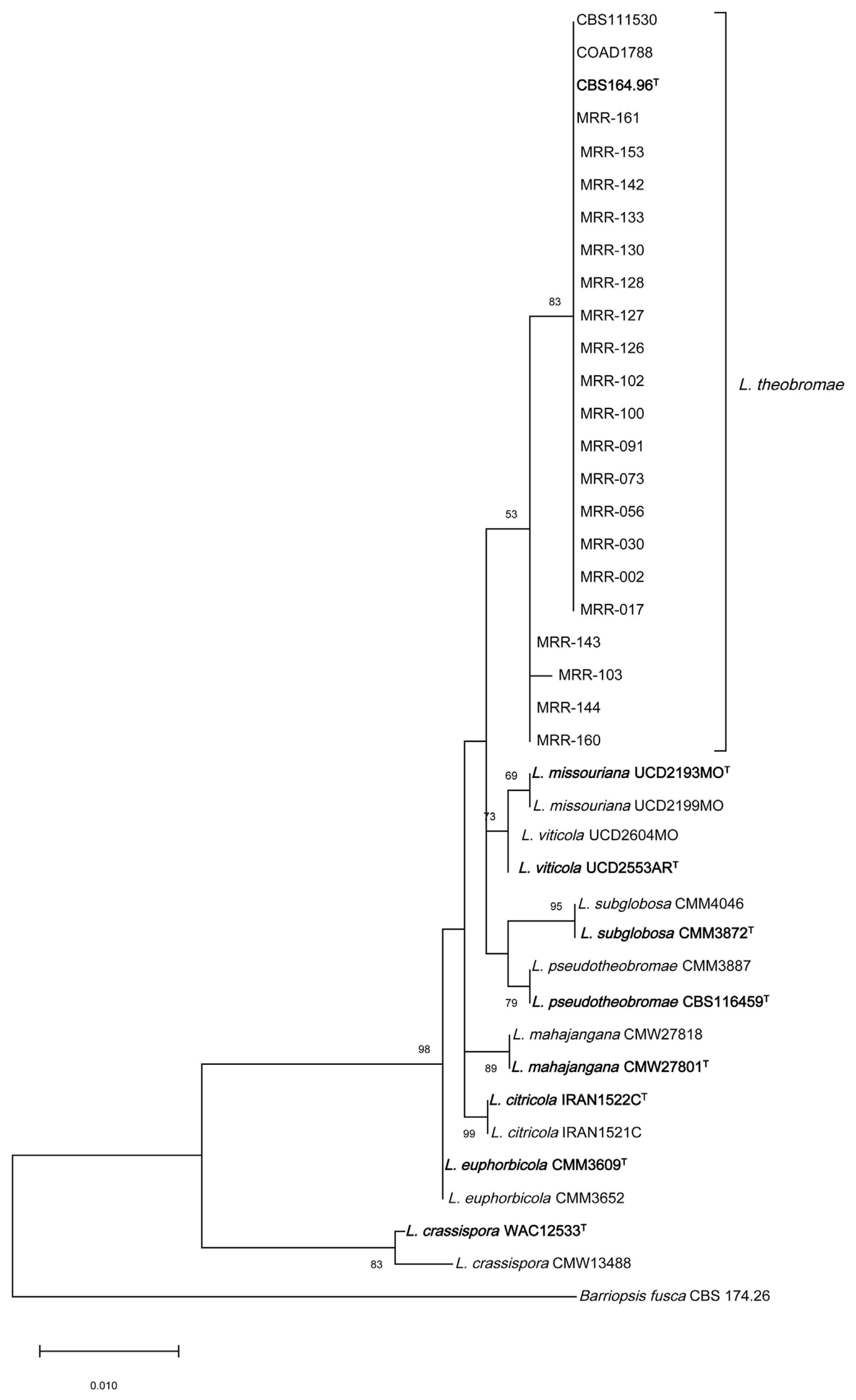
Twenty L. theobromae isolates were selected for phylogenetic analysis using ITS and β-tubulin regions based on the geographical region, representing the four states of South India. Also, based on virulence and morphological characters of the isolates like colony color, characters of aerial mycelia growth. The amplicon size of ITS and β-tubulin was detected between 500-600 bp and 300-400 bp, respectively (Munirah et al., 2017). Blast search homology of ITS and β-tubulin sequences revealed 98-100% similarity with type strain CBS 164.96 L. theobromae (Supplementary Table 2). GenBank accession numbers were obtained for all the isolates (Table 3). A phylogenetic tree was constructed for all the isolates of L. theobromae and the isolates were grouped into a single clade, representing the reference sequences (CBS111530 and COAD1788) and type strain (CBS 164.96) with a bootstrap value of 83% (Fig. 4). Thus confirming the identity of the isolated pathogen is L. theobromae, whereas all the other Lasiodiplodia species; L. citricola, L. crassispora, L. euphorbicola, L. mahajangana; L. missouriana; L. pseudotheobromae; L. subglobosa formed monophyletic clade individually with respect to their type strain. The closest species near to L. theobromae isolates were L. missouriana and L. viticola, whereas L. crassispora formed a separate cluster. The final pooled dataset of ITS and β-tubulin of 40 nucleotide sequences after alignment consists of 649 positions, 595 were conserved, 54 variable, 31 parsimony-informative, and 23 were singleton.
Pathogenicity test
The pathogenicity test conducted on susceptible genotypes Victory-1 and Thailand Male revealed significant variation (P < 0.05) between the selected 20 isolates (Table 4). The infected plants, showed similar symptoms of BRR, like the appearance of dark colored lesions, rotted roots, wilting of leaves and browning of stems (Fig. 5). The wilting and rotting percentage of Victory-1 ranged between 3.6 to 97.6 and 4.6 to 100.0, respectively, similarly of Thailand male, 7.6 to 99.3 and 10.3 to 95.0, respectively. Sixteen isolates were highly pathogenic and four isolates, MRR-056, MRR-126, MRR-133, and MRR-153 were less pathogenic on both the susceptible genotypes exhibiting low levels of wilting and rotting. Among all the isolates, MRR-142, isolated from Andhra Pradesh, was found to be highly pathogenic which caused a maximum percentage of wilting and rotting (Table 4).
Screening of resistant mulberry genotypes
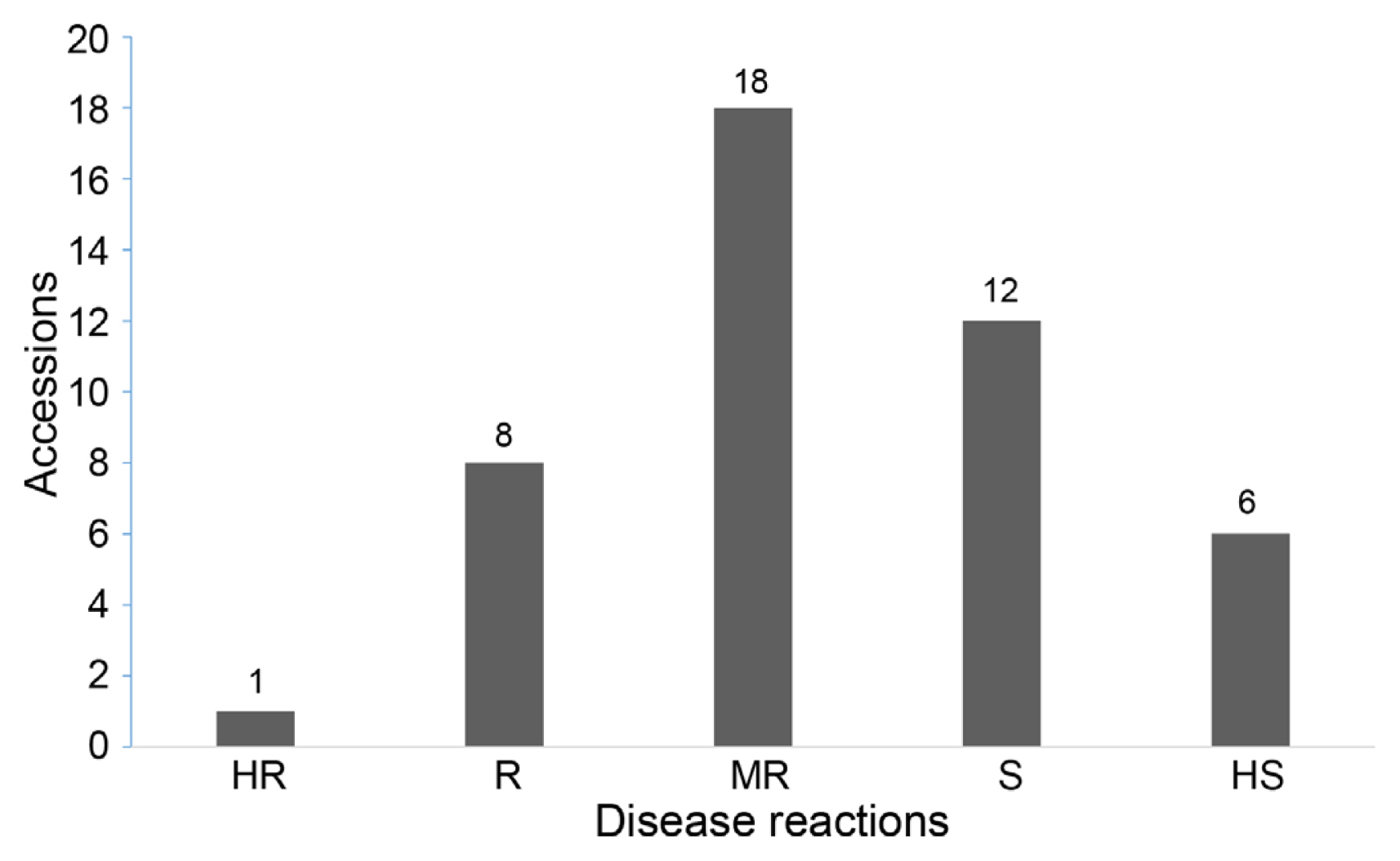
A total of 45 mulberry accessions were screened against the L. theobromae isolate MRR-142 through the root dip method of inoculation. The BRR disease reactions of resistant and susceptible accessions revealed significant variation under glasshouse conditions. The wilting and rotting percentage among the 45 accessions in the two experiments ranged between 0.0 to 96.7 and 0.0 to 100.0, respectively (Table 5). Accession, ME-0168 of Indonesian origin belonging to M. latifolia was found to be highly resistant (HR) against BRR (Fig. 6). The exotic accession ME-0168 was free from BRR symptoms and was on par with healthy control plants. Eight accessions that were found to be resistant (R) were G2, ME-0006, ME-0011, ME-0093, MI-0006, MI-0291, MI-0489 and MI-0501. Whereas six accessions showed highly susceptible (HS) reaction, 12 were susceptible, and 18 were moderately resistant (MR) to BRR (Table 5, Fig. 7).
Discussion
Mulberry is affected by a variety of diseases produced by fungi and bacteria leading to severe yield loss and reducing the nutritive value of leaves. The repeated harvesting of leaves during the cultivation of mulberry leads to the depletion of soil nutrients and makes plants vulnerable to soil-borne diseases (Narayanan et al., 2015). Among the many fungal pathogens involved in causing root rot disease of mulberry, L. theobromae is a dominating fungus causing severe yield loss due to a decrease in plant population. L. theobromae has a broad range of plant hosts and is more prevalent in tropical and subtropical countries. Although several studies reported the pathogenic potential of this fungal species, it is also capable of surviving and spreading as an endophytic plant associate (de Silva et al., 2019; Muniz et al., 2011; Salvatore et al., 2020).
The study of micromorphological and cultural features is indispensable for the characterization of fungal isolates. In the case of genus Lasiodiplodia microscopic examination of conidia and paraphyses is important as these features differentiate various species of this genus (Rosado et al., 2016). Like several other research investigations, the present study also utilized SEM observations for interpreting micromorphological features of 20 L. theobromae isolates (Alves et al., 2008; Muniz et al., 2011; Tovar-Pedraza et al., 2012). All the sporulating isolates showed pycnidia with septate paraphyses, which is a characteristic feature of L. theobromae (Latha et al., 2013). The mature conidia appeared dark brown with thick wall and longitudinal striations. The mature conidial length was found in the range from 25 μm to 17 (± 3.0) μm and breadth was in the range from 14 μm to 7 (± 2.0) μm. PDA was frequently used to study the growth characteristics of L. theobromae isolates as this medium composition favored maximum growth of L. theobromae (Latha et al., 2013). L. theobromae isolates grew as irregular shaped colonies with greenish black/grey color and black color in reverse (Pečenka et al., 2021).
Based on cultural and morphological features, the isolated fungal pathogens were identified as L. theobromae. Twenty selected isolates representing four states of South India were subjected to prove Koch’s postulates which confirmed their pathogenicity with varying degrees of disease severity. However, the presence of highly varied cultural and morphological characteristics, diverse host range, ability to cause different symptoms and widespread geographical distribution of L. theobromae alarms about the existence of several strains in nature (Slippers et al., 2013; Sowmya et al., 2018; Xie et al., 2014). The molecular classification is rapid and provides precise phylogenetic distinctiveness.
Phylogenetic analysis of ITS and β-tubulin regions confirmed the identity of the selected isolates as L. theobromae. The 20 isolates of L. theobromae from this study, type strain (CBS 164.96) and reference sequences (CBS 111530 and COAD 1788) were grouped into a single cluster and separated from other known species of Lasiodiplodia. Even though the isolates were grouped into a single clade, four isolates (MRR-103, 143, 144, and 160) were away from the type strain (Fig. 4), the bootstrap values were low (53%) and the sequence homology of these four isolates was 98% as compared to the other isolates which had a high similarity of 100% with the type strain with high bootstrap values (83%). Likewise, Pappachan et al. (2020), isolated L. theobromae isolate from the infected roots of mulberry collected from Mizoram, Northeast India has 99.03% homologous to L. theobromae Bl16 (MK813947) with low bootstrap value.
Sowmya et al. (2018) based on cultural, morphological, pathogenicity, SSR and RAPD markers revealed significant variations among the L. theobromae isolates obtained from the infected gardens of Karnataka, Andhra Pradesh and Tamil Nadu. Also from her study, she could not establish a clear correlation between genetic diversity and geographical distribution. Many researchers in other crops, identified different species of Lasiodiplodia, however, from our studies, we could only identify L. theobromae causing BRR of mulberry in India (Pappachan et al., 2020; Radhakrishnan et al., 1995; Sowmya et al., 2018; Sukumar and Padma 1999) and this is the first detailed phylogenetic analysis of L. theobromae causing BRR of mulberry in India using two genes.
The wild species of mulberry (M. serrata and M. laevigata) are known to possess several important resistant genes to many biotic and abiotic factors (Tikader and Dandin, 2007; Vijayan et al., 2011). Even though 68 species from Morus were available, only a limited number of species have been utilized in developing mulberry varieties (Datta, 2000). The emerging new varieties from the narrow gene pool are more homogenous and become HS to pathogens and vulnerable to poor environmental conditions (Vijayan et al., 2011). Cultivation of disease resistant varieties is one of the best approaches to exclude plant pathogens. The selection of promising genotypes will contribute to developing resistant breeding methods.
Screening of mulberry germplasm using pathogenic L. theobromae isolates, and identification of highly durable BRR resistant varieties can aid in achieving sustainable sericulture. Mulberry researchers in India have identified resistant accessions to bacterial leaf spot (Xanthomonas campestris), powdery mildew (Phyllactinia corylea), charcoal root rot (M. phaseolina), and root knot (Arunakumar et al., 2021; Banerjee et al., 2009; Chattopadhyay et al., 2010; Maji, 2011; Pinto et al., 2018). To our knowledge, there is no report on screening of germplasm resistance against L. theobromae causing BRR. To select promising mulberry germplasm against BRR, the present study attempted to evaluate 45 diverse accessions following the root dip method of inoculation under glasshouse conditions. In the repeated experiments, the mulberry accession ME-0168 which is an Indonesian origin was consistently found to be HR to BRR. Other eight accessions, G2, ME-0006, ME-0011, ME-0093, MI-0006, MI-0291, MI-0489, and MI-0501 were found to be resistant.
Most of the resistant accessions, ME-0006, ME-0011, ME-0093, including HR (ME-0168) and MR (ME-0066 and ME-0141) were from exotic collections and belonged to M. latifolia. This indicates M. latifolia possesses resistant genes against BRR of mulberry. The other four resistant accessions, MI-0006, MI-0291, MI-0489, and MI-0501, belonged to M. indica. Joty et al. (2019) found that the leaf extracts of M. indica and M. latifolia induced the maximum antibacterial and antioxidant activity against the pathogenic bacteria. Interestingly, G2 high yielding mulberry variety was found to be resistant to BRR is a hybrid of ME-0168 and S-34, similarly, G4 which is MR to BRR is a cross pollinated hybrid of ME-0168 and S-30. This proves that the identified HR accession ME-0168 along with the other promising resistant sources can be exploited in mulberry breeding for developing BRR resistant varieties and to develop mapping populations which successively helps in the identification of molecular markers associated with BRR.
In conclusion, the outcomes of this research represent the detailed study of phylogeny and pathogenicity of L. theobromae causing BRR of mulberry in India. The MRR-142 isolate, isolated from Ananthapur, Andhra Pradesh was found to be highly pathogenic which caused a maximum percentage of wilting and rotting. Further, the same isolate was used to screen resistant accessions. The root dip inoculation method identified the HR accession ME-0168 against BRR of mulberry. ME-0168, along with the other sources of resistance can be utilized as resistant parents for developing BRR resistance varieties.




 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement1
Supplement1 Print
Print






