Identification of Endogenous Genes for Normalizing Titer Variation of Citrus Tristeza Virus in Aphids at Different Post-acquisition Feeding Times
Article information
Abstract
Citrus tristeza virus (CTV) is efficiently transmitted in a semi-persistent manner by the brown citrus aphid (Toxoptera citricida (Kirkaldy)). Currently, the most sensitive method for detecting plant viruses in insect vectors is reverse-transcription quantitative polymerase chain reaction (RT-qPCR). In this study, the elongation factor-1 alpha (EF-1α) gene and acidic p0 ribosomal protein (RPAP0) gene were confirmed to be suitable reference genes for RT-qPCR normalization in viruliferous T. citricida aphids using the geNorm, NormFinder, and BestKeeper tools. Then the relative CTV titer in aphids (T. citricida) at different post-acquisition feeding times on healthy plants was quantified by RT-qPCR using EF-1α and RPAP0 as reference genes. The relative CTV titer retained in the aphids gradually decreased with increasing feeding time. During the first 0.5 h of feeding time on healthy plants, the remaining CTV titer in aphids showed about 80% rapid loss for the highly transmissible isolate CT11A and 40% loss for the poorly transmissible isolate CTLJ. The relative CTV titer in aphids during more than 12 h post-acquisition times for CT11A was significantly lower than at the other feeding times, which is similar to the trend found for CTLJ. To our knowledge, this is the first report about the relative titer variation of CTV remaining in T. citricida at different post-acquisition feeding times on healthy plants.
Citrus tristeza virus (CTV), a member of the genus Closterovirus, is a serious threat to citrus production (Bar-Joseph et al., 1989; Hilf et al., 2005), and brown citrus aphids (Toxoptera citricida (Kirkaldy)) are the most efficient vector of CTV (Moreno et al., 2008; Rocha-Peña et al., 1995). There are many factors influencing CTV transmission mediated by its aphid vectors, such as the CTV isolate genotype, environmental conditions, and acquisition or infection feeding periods (Roistacher and Bar-Joseph, 1987; Sharma, 1989; Zhou et al., 2011). Toxoptera citricida has a near-maximal transmission efficiency 4 h after acquisition feeding (Costa and Grant, 1951). However, a 6-h acquisition or infection feeding period is required for Aphis gossypii to obtain near-maximal transmission (Raccah et al., 1976). Moreover, A. gossypii maintains low transmissibility after feeding for 6 h on an intermediate lime seedling and has no transmissibility for 24 h (Raccah et al., 1976). Based on polymerase chain reaction (PCR) assays, CTV has been detected in several aphid species, such as T. citricida, A. gossypii, and T. aurantii (Mehta et al., 1997; Olmos et al., 1999). In addition, reverse-transcription quantitative polymerase chain reaction (RT-qPCR), which is more sensitive than conventional PCR, has been reported for the exact quantification of CTV in A. gossypii and T. citricida (Bertolini et al., 2008; Liu et al., 2019; Saponari et al., 2008). The quantity of CTV acquired by T. citricida aphids on plants gradually increased and then decreased 4–6 h after the acquisition access periods (Liu et al., 2019). In further experiments, the transmission rates were positively associated with the viral titer in the vector in the first 6 h of the acquisition access periods (Liu et al., 2019). Although other previous studies measured the virus titer differences, they did not use validated reference genes. However, there are very few studies about the titer changes of virus particles remaining in aphids at different post-acquisition feeding times on healthy plants.
RT-qPCR is a powerful tool for quantifying the virus in biological samples after normalization, using endogenous genes as internal references (Bertin et al., 2016; Maroniche et al., 2011). For example, translation elongation factor 1α (EF-1α) gene and Actin1 gene are used as internal reference genes to normalize virus titer in yellow dwarf viruses viruliferous aphids (Wu et al., 2014). It is very important to identify suitable reference gene(s) for different experiments, as the transcript levels of reference genes vary with experimental conditions (Zhang et al., 2018). Thus far, no research has been found on the transcript changes in aphids feeding on CTV-infected citrus, and no appropriate internal reference genes have been identified in viruliferous T. citricida aphids. Although no T. citricida aphid genome sequencing project has been announced, great progress has been made in expressed sequence tag sequences from various tissues of T. citricida aphids (Hunter et al., 2003), which will enable the discovery of the molecular mechanism for virus transmission by insects.
In this study, the relative titer variation of CTV in aphids was evaluated at different post-acquisition feeding times on healthy plants. To identify genes suitable for the normalization of virus titer, three common algorithms, geNorm, NormFinder, and BestKeeper, were first performed, and the expression stability of the five candidate reference genes (RPL5, RPAP0, EF-1α, EF-2, and M-Act) of CTV-viruliferous T. citricida was compared.
Materials and Methods
Virus source
Two mild CTV isolates, CT11A and CTLJ, were used in this study. The single-aphid transmission rates of CT11A and CTLJ were 65.6% and 4.4%, respectively (Liu et al., 2019). Both CTV isolates were collected from ‘Jincheng’ sweet orange (Citrus sinensis (L.)) at the Citrus Research Institute, Southwest University/Chinese Academy of Agricultural Sciences. The viruses were then graft-inoculated onto ‘Jincheng’ sweet orange seedlings for conservation.
Insect culture
CTV-free T. citricida colonies were established as described by Broadbent et al. (1996) and reared on sweet orange seedlings in insectaries at 25°C and a photoperiod of 16 h light/8 h dark.
Sample preparation and RNA extraction
To identify the stability of reference genes, hundreds of the apterous adult aphids were fed for 6 h on sweet orange seedlings infected with CT11A or CTLJ isolates, based on the previous observation that the maximum CTV titration in aphids was at the acquisition access period of 4–6 h (Liu et al., 2019). The aphids were randomly collected from CTV-infected and healthy sweet orange seedlings with 15 T. citricida aphids per group. This experiment was repeated three times.
To assess the CTV titer variation in aphids at different post-acquisition feeding times, thousands of non-viruliferous aphids, which had been apterous adults of 1–2 days old, were fed on CTV-infected ‘Jincheng’ sweet orange seedlings. After a 6-h acquisition access period on CTV-infected plants, 45 adult aphids were collected for virus detection, and 450 adult aphids were used for post-acquisition feeding on healthy ‘Jincheng’ sweet orange seedlings for each replication. Ten sampling events (0.5, 1, 2, 3, 4, 6, 9, 12, 24, and 48 h) were performed during feeding time on healthy sweet orange seedlings. At the completion of each feeding period, total RNA was extracted, and CTV was quantified by RT-qPCR in three groups, with 15 aphids per group. EF-1α and RPAP0 were used as reference genes in viruliferous T. citricida. Individuals in each group were randomly collected from several sweet orange plants during each feeding period. This experiment was repeated three times.
The total RNA of adult aphid samples was extracted with a TransZol Up Kit (TransGen Biotech, Beijing, China) following the manufacturer’s protocol (Liu et al., 2019). The total RNA was resuspended in 20 μl of RNase-free water.
Reverse transcription and qPCR assay
In this study, primer HD-F (5′-CGAGTCTGGGGTAGAAGTAACGA-3′) paired with primer HD-R (5′-GGTCAAGAATCTGCACACAGG-3′) was used to amplify a specific 136-bp nucleotide sequence of the coat protein gene from CTV (Liu et al., 2019).
The five candidate reference genes selected for this study were genes encoding ribosomal proteins (RPL5, RPAP0), elongation factors 1α and 2 (EF-1α, EF-2), and muscle actin (M-Act), all of which are publicly available insect reference genes (Li et al., 2013; Maroniche et al., 2011; Zhang et al., 2018). The full-length sequences of the five genes were downloaded from the NCBI database (https://www.ncbi.nlm.nih.gov/), and primers were designed for RT-qPCR using Beacon Designer 7 (Premier Biosoft, Palo Alto, CA, USA). Their nomenclature, such as names, accession numbers, description, and the designed RT-qPCR primers, are presented in Table 1.

Candidate reference genes and their primer sequences used in this study
cDNA was synthesized from T. citricida aphids’ RNA using the TransScript One-Step gDNA remover and cDNA Synthesis Kit (TransGen Biotech, Beijing, China) according to the manufacturer’s instructions. The reaction volume was 10 μl, containing 500 ng of RNA, 0.5 μM of reverse primer, 5 μl of reverse transcription buffer, 0.5 μl of PrimerScript TM/RI Enzyme Mix, 0.5 μl of gDNA Remover, and RNase-free H2O. The reverse transcription reaction was performed on a C1000 Thermal Cycler (Bio-Rad, Hercules, CA, USA), and the procedure was as follows: 42°C for 30 min and 85°C for 5 s. The resulting cDNA was stored at −20°C until use.
Real-time PCR was performed in a 20-μl reaction volume that contained 1× SYBR Premix ExTaq II enzyme (Takara, Dalian, China), 0.6 μM of each gene-specific primer, and 2 μl of the first strand of cDNA. The PCR reaction procedure was as follows: denaturation at 95°C for 30 s, 40 cycles of denaturation at 95°C for 10 s, annealing at 60°C for 15 s, and extension at 72°C for 20 s. Once PCR was finished, the PCR reaction product was incubated at 95°C for 15 s, annealed at 65°C for 30 s, and gradually increased to 95°C again for melting curve analysis. Data acquisition and analysis were performed using the iCycleriQ Real-Time PCR Detection System (Bio-Rad).
Data analysis and statistical analysis
Three Microsoft Excel-based common software tools, geNorm (Vandesompele et al., 2002), Normfinder (Andersen et al., 2004), and BestKeeper (Pfaffl et al., 2004), were used to explore gene expression stability. The optimum internal reference gene number was estimated using the geNorm pairwise variation value (vn/n+1) between two sequential normalization factors (Vandesompele et al., 2002).
The relative CTV titer variation in the aphids was calculated using the comparative CT method (2−ΔΔCT method) by normalizing to two reference genes, EF-1α and RPAP0 (Livak and Schmittgen, 2001). Using the statistics software SPSS ver. 12.0 (SPSS Inc., Chicago, IL, USA), one-way analysis of variance (ANOVA) and Duncan’s multiple comparisons (α = 0.05) was run to measure the relative titer variation in CTV levels in T. citricida.
Results
Identification of candidate reference genes
Five reference genes were used for RT-qPCR normalization. RT-qPCR primers efficiently amplified the desired sequences of five common candidate genes from CTV-viruliferous aphids. The amplification efficiency of these five pairs of RT-qPCR primers ranged from 94.7% for the EF-1α gene to 105.2% for the M-Act gene. Correlation coefficients (R2) ranged from 0.991 for RPL5 gene to 0.998 for EF-1α gene (Table 1). A single peak in the melting curve of each candidate reference gene was shown using iCycleriQ software. The study focused on EF-1α gene and RPAP0 gene. Fig. 1 shows their melting curves.
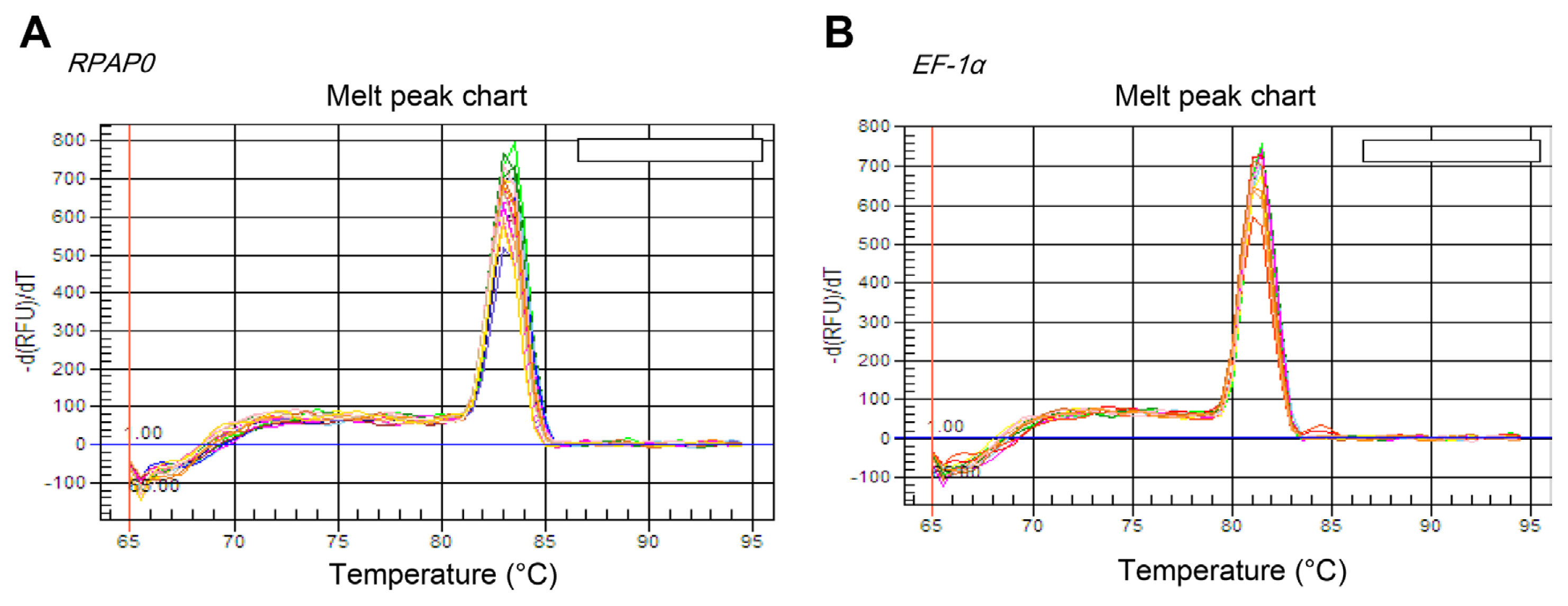
The melting curve for RPAP0 (A) and EF-1α (B) with single peaks obtained from three technical replicates. Real-time polymerase chain reaction (PCR) was performed in a 20-μl reaction volume that contained 1× SYBR Premix ExTaq II enzyme, 0.6 μM of each gene-specific primer, and 2 μl of the first strand of cDNA. The PCR reaction procedure was as follows: denaturation at 95°C for 30 s, 40 cycles of denaturation at 95°C for 10 s, annealing at 60°C for 15 s, and extension at 72°C for 20 s. Once PCR was finished, the PCR reaction product was incubated at 95°C for 15 s, annealed at 65°C for 30 s, and gradually increased to 95°C again for melting curve analysis. Data acquisition and analysis were performed using the iCycleriQ Real-Time PCR Detection System.
Expression stability of candidate reference genes
Based on the average pairwise variation of each candidate reference gene compared to the other four genes, geNorm assigns a stability index M to the gene, and a low M-value indicates high gene stability (Vandesompele et al., 2002). RPAP0 gene showed the lowest M-value among the five selected genes; thus, RPAP0 gene was the most stable of these candidate reference genes. EF-1α gene was the second most stable reference gene, while the other three genes (EF-2, M-Act, and RPL5) were not very stable in CTV-viruliferous aphids (Table 2). In addition, geNorm revealed that RPAP0/EF-1α gene was the most stable reference gene pair (Fig. 2). Normfinder focuses on the inter- and intra-group variations and calculates the stability values (SVs) of all candidate reference genes. The lower the SV of a gene, the more stable its expression (Andersen et al., 2004). Normfinder also identified RPAP0 gene as the most stable gene (SV = 0.14). The stability of the five candidate genes was ranked as follows: RPAP0 > EF-1α > EF-2 > M-Act > RPL5 (Table 2). BestKeeper is a tool for calculating the standard deviation of the Ct value of genes and the coefficient of variance (CV) between genes, thus indicating gene stability (Pfaffl et al., 2004). A gene showed more stability when the CV value became smaller. BestKeeper analysis demonstrated that RPAP0 gene ranked as the least variable gene with a CV of 1.74, followed by EF-1α gene and EF-2 gene, while M-Act and RPL5 were the most variable genes (Table 2). In summary, geNorm, NormFinder, and BestKeeper showed consistent results, and RPAP0, EF-1α, and EF-2 were the three most stable genes of the five candidate reference genes in CTV-viruliferous aphids (Table 2).
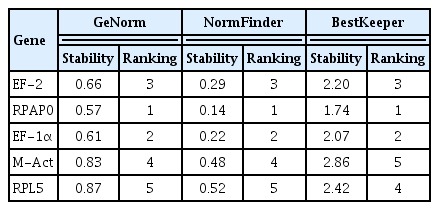
Expression stability of reference genes in viruliferous Toxoptera citricida analyzed by geNorm, NormFinder, and BestKeeper
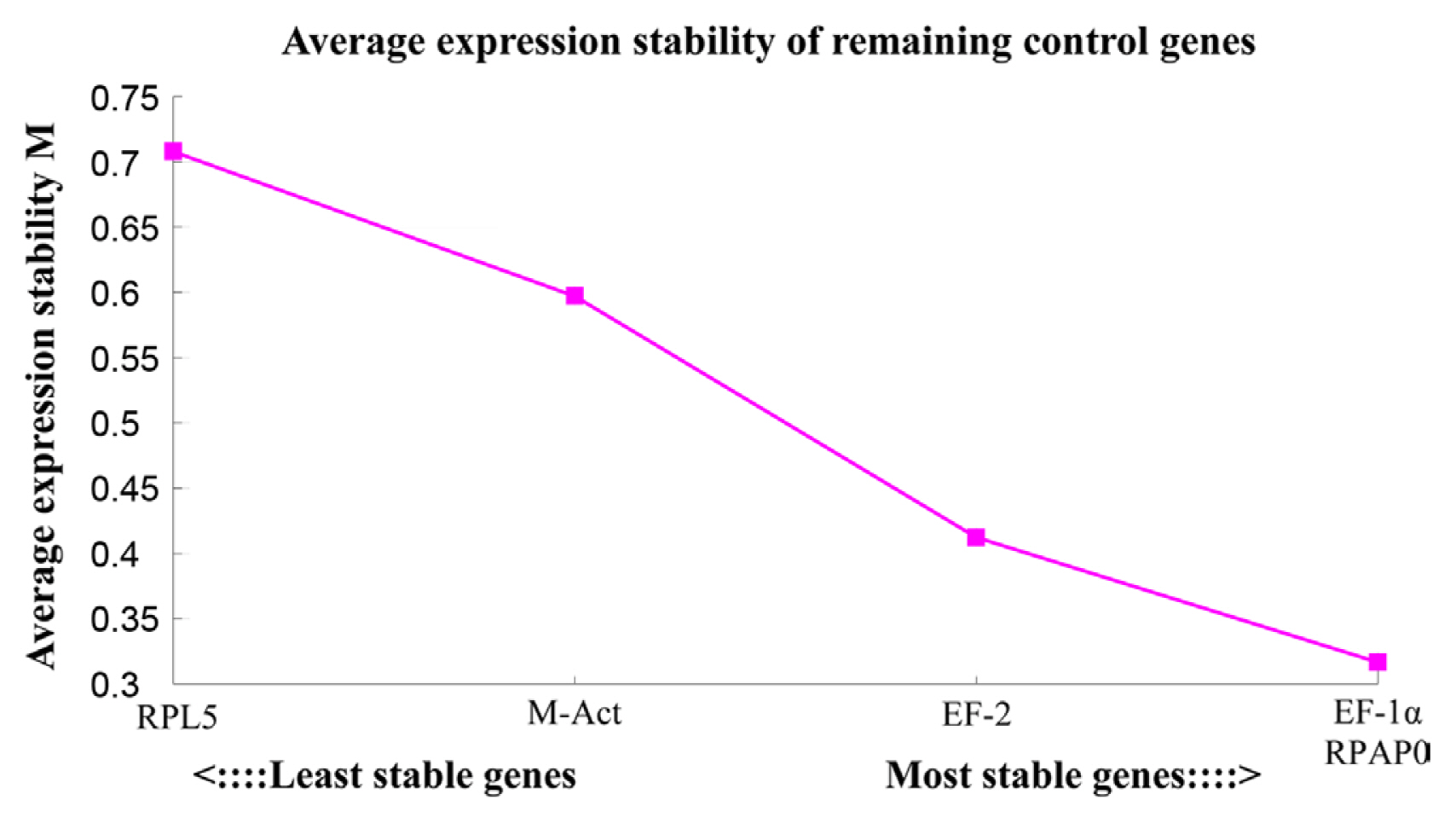
Average expression stability value of candidate reference genes in viruliferous Toxoptera citricida analyzed by geNorm. Based on the average pairwise variation of each candidate reference gene compared to the other four genes, the geNorm assigns a stability index M to the gene, and then all the studied genes are ranked. The least stable gene with the highest M-value is excluded sequentially (from left to right) until the most stable gene pair remains.
The number of reference genes for RT-qPCR data normalization can be determined by pairwise variations (vn/n+1) based on geNorm software. A pairwise variation (vn/n+1) below 0.15 indicates that no additional genes are required for the calculation of the normalization factor NFn (Vandesompele et al., 2002). GeNorm showed that the pairwise variation v2/3 of 0.144 was below the proposed threshold value of 0.15 (Vandesompele et al., 2002), while the values of v3/4 and v4/5 were higher than 0.15 (Fig. 3), indicating that RPAP0 gene and EF-1α gene should be selected together as reference genes for RT-qPCR normalization in our experimental conditions.

Optimal number of reference genes for normalization in viruliferous Toxoptera citricida by geNorm. The pairwise variation (Vn/n+1) analysis showing the normalization factor (NF) variability (V) between the NFn and NFn+1 that results from including an additional gene in the NFn calculation. Pairwise variations (vn/n+1) under 0.15 indicate that no additional genes are required for normalization.
Virus titer variations at different post-acquisition feeding times
Using RPAP0 gene and EF-1α geneas multiple reference genes, the relative amount of CTV in aphids was evaluated after they had undergone different post-acquisition feeding times on healthy sweet orange seedlings (Fig. 4). For the highly transmissible isolate CT11A, the remaining CTV in aphids rapidly decreased, changing from 2.58 to 0.50, during the post-acquisition feeding time between 0 and 0.5 h, indicating a loss of about 80%. The virus level then declined at a decreased rate from 1 to 48 h. Similar trends were displayed for the relative amount of the poorly transmissible isolate CTLJ in the aphids during 0.5–48 h of post-acquisition time on healthy sweet orange seedlings, although with different kinetics from 0 to 3 h. The remaining CTLJ amount in the aphids showed a loss of about 40% (1.83-1.09) during the first 0.5 h of feeding time and reached about 92% (1.83-0.14) during the 0–3-h post-acquisition feeding time (Fig. 4). At the 3-h post-acquisition feeding time, the relative quantification of CTV in aphids was similar for CT11A and CTLJ isolates. The relative CTV titers in aphids during the post-acquisition time of 3–48 h for CTLJ were significantly lower than during 0.5–2 h, which is similar to the trend found for CT11A. The relative CTV titers in aphids during the post-acquisition time of 12–48 h were significantly lower than those at the other feeding times, excluding at 9 h for CTLJ (Fig. 4).
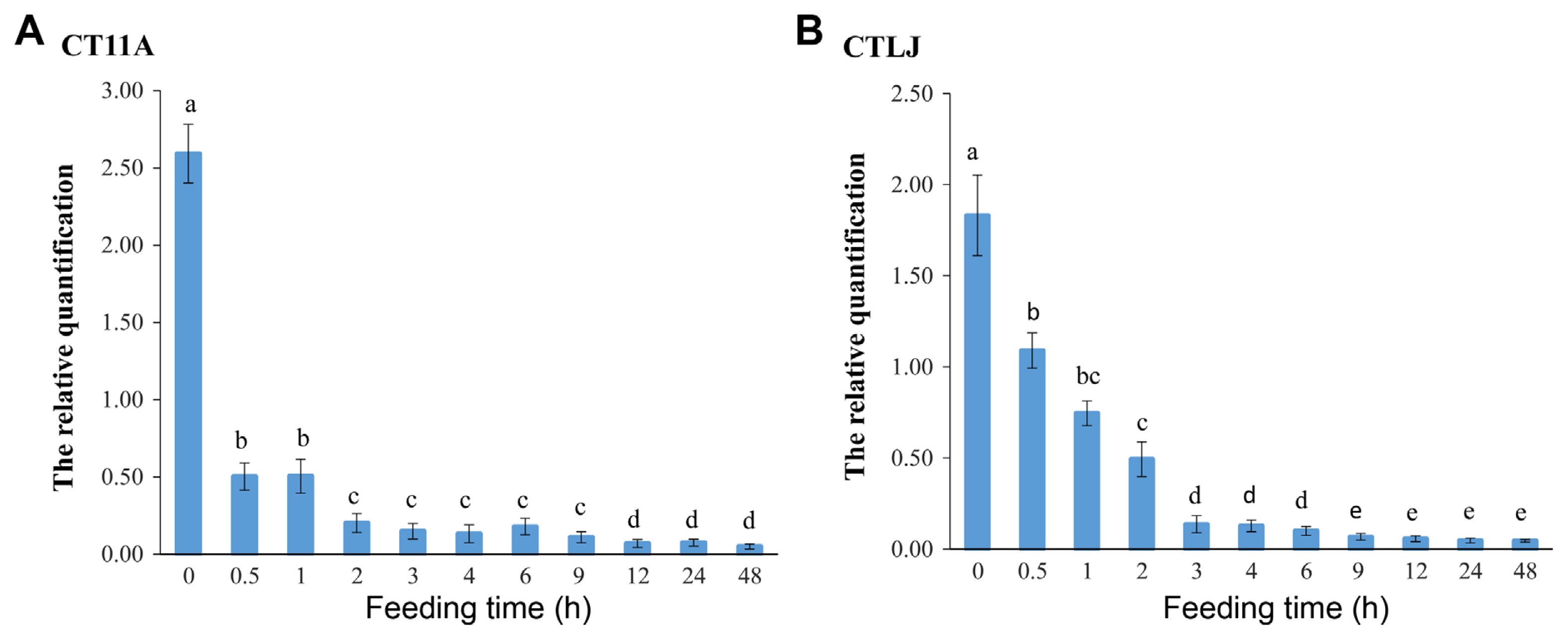
Citrus tristeza virus (CTV) titer variation in Toxoptera citricida after different post-acquisition feeding times on healthy plants. Thousands of non-viruliferous aphids, which had been apterous adults of 1–2 days old, were fed on CTV-infected ‘Jincheng’ sweet orange seedlings. After a 6 h acquisition access period on CTV-infected ‘Jincheng’ sweet orange plants, 45 adult aphids were collected for virus detection, and 450 adult aphids were used for post-acquisition feeding on healthy ‘Jincheng’ sweet orange seedling for each replication. Ten sampling events (0.5, 1, 2, 3, 4, 6, 9, 12, 24, and 48 h) were performed during feeding time on healthy sweet orange seedlings. This experiment was repeated three times. At the completion of each feeding period, total RNA was extracted from three groups of 15 aphids, and CTV was quantified by reverse-transcription quantitative polymerase chain reaction in T. citricida. Individuals in each group were randomly collected from several sweet orange plants during each feeding period. EF-1α and RPAP0 were used as reference genes in viruliferous T. citricida. Duncan’s multiple comparison was explored. The same letter indicates no significant difference at P < 0.05.
Discussion
To understand the mechanistic basis of insect-virus interactions, a reliable quantitation method is necessary. Moreover, to eliminate the influence of RNA quality and the reverse transcription efficiency of different samples on the results, appropriate reference genes are needed for RT-qPCR analysis (Mason et al., 2008; Xie et al., 2019). Although there are many reports on reference genes remaining stable under various experimental conditions, no reference genes are stable under all biological and abiotic factors (Gutierrez et al., 2008; Kim and Yun, 2011). Thus, two or more reference genes used for normalizing RT-PCR results under different experimental conditions are essential. In this study, three common methods of reference gene analysis, geNorm, NormFinder, and BestKeeper, for five candidate reference genes in CTV-viruliferous aphids were used, and two genes (RPAP0 and EF-1α) of them were targeted for normalizing gene expression in viruliferous aphids. Generally, actin gene is used as the reference gene in RT-qPCR experiments and is stably expressed in mal de río cuarto virus-infective planthoppers (Maroniche et al., 2011). In our study, however, its performance was unsatisfactory, meaning that a common reference gene might not be the optimal choice for some studies. EF-1a gene is a stable gene in T. citricida aphids under starvation and UV irradiation stress conditions (Shang et al., 2015). Similarly, in our experimental conditions, EF-1α gene was also a stable reference gene. Moreover, EF-1α gene is a stably expressed reference gene for RT-qPCR experiments in many insects, such as Spodoptera litura (Lu et al., 2013) and Bemisia tabaci (Li et al., 2013), but it is variable in viruliferous planthoppers (Maroniche et al., 2011). It is evident that the stability of the reference gene depends on insect species and experimental conditions (Liang et al., 2014).
Many studies have estimated the number of viral particles carried in insect vectors (Betancourt et al., 2008; Moreno et al., 2009; Moury et al., 2007). However, very few studies have been conducted on virus quantification in vectors at different post-acquisition feeding times on healthy plants. In this study, analysis of the remaining CTV titer in aphids showed a loss of about 80% for CT11A, while a loss of about 40% for CTLJ was observed during the first 0.5 h of feeding time on healthy plants (Fig. 4). In previous studies, the single-aphid transmission rates of CT11A and CTLJ were 65.6% and 4.4%, respectively (Liu et al., 2019). According to Raccah et al. (1980), CT11A and CTLJ belong to the highly (over 21%) and poorly (less than 5%) transmissible isolates, respectively. The present results indicate that aphids acquiring highly transmissible isolates release virions more quickly than aphids acquiring less transmissible isolates. Moreover, the relative quantification of CTV was higher in aphids fed on plants infected with CT11A (2.58) than with CTLJ (1.83) at the beginning of the post-acquisition feeding time (Fig. 4), which is similar to the results that aphids acquired more virions of the highly transmissible isolate (Liu et al., 2019; Zhou et al., 2011). These results indicate that more virions of the highly transmissible isolate are not only acquired by aphids, but are also quickly released compared to those of the lowly transmissible isolate. The interactions between viruses and their vectors are highly specific. The coat protein and helper component proteins of plant viruses have been reported to be involved in several virus–vector interactions (Blanc et al., 1998; Brault et al., 1995). Mutations of these proteins have been found to reduce or abolish viral transmission by insect vector (Stewart et al., 2010). Thus far, the mechanisms for the release of virions from the vector into plants have not been well explained. It may be determined by specific vector proteins and virus-encoded proteins (Ng and Falk, 2006; Uzest et al., 2007). Killiny et al. (2016) found that CTV virions bound specifically to the cuticular surface of the aphid cibarium, and three virally-encoded proteins (p27, p65, and p61) may be involved in vector interaction. Sequence variations in the p65 and p61 genes determine the efficiency of CTV transmission by the brown citrus aphid (Harper et al., 2016). Some mutations of p27 gene nucleotide sequence of nontransmissible isolates led to substitution of new amino acids with a different polarity and charge (Barzegar et al., 2010). There are nine amino acid mutations in the p27 gene deduced amino acid sequence of the poorly transmissible CTLJ isolate (Liu et al., 2021). Moreover, the full genome sequence of CT11A is distantly related to the poorly transmissible isolate T36 (Liu et al., 2019). This implies that sequence variation in CTV isolates can affect the release of CTV. In addition, the quantity of CTV in aphids particularly depends on insect vector feeding behavior (Jiang et al., 2000). Limburg et al. (1997) reported that an average of 20.5 min was required to reach the phloem sieve elements for the stylets of A. fabae, which is the insect vector of beet yellow virus (BYV), another Closterovirus. This was consistent with the minimum time period of 20 min for BYV acquisition access (Limburg et al., 1997). For CTV, about 30 min is required to reach the phloem position for T. citricida because CTV can be acquired by T. citricida at low levels in 30 min (Bar-Joseph et al., 1989; Liu et al., 2019; Raccah et al., 1976). This implies that the quantity of CTV in aphids showed a rapid decrease during the phloem ingestion phase within 30 min. In addition, the remaining CTLJ amount in the aphids showed a loss of more than 92% during the 0–3-h post-acquisition feeding time (Fig. 4). However, the near-maximal transmission efficiency of CTV was reached after 6 h of the infection feeding period (Raccah et al., 1976). This indicates that a longer infection feeding time is still needed to release CTV retention on the cibarium to attain near-maximal transmission rates, although the CTV targets rapidly decrease in aphids during 3 h of feeding on healthy plants.
Before RT-qPCR was developed, the retention of viruses was evaluated according to the length of time that the insect vector retained transmissibility under post-acquisition feeding conditions (Raccah et al., 1976). Raccah et al. (1976) showed that A. gossypii aphids lost CTV transmissibility at 24 h post-acquisition (Raccah et al., 1976). In this study, the relative CTV titer in aphids during more than 12 h post-acquisition times was significantly lower than those during 0.5–9 h feeding times on healthy plants (Fig. 4). These results support the findings of Raccah et al. (1976). In addition, for BYV, which is also transmitted in a semi-persistent manner, the transmissibility by A. fabae decreased by more than 50% during the first 3 h post-acquisition feeding period, and the decrease was more gradual from 3 to 48 h (Limburg et al., 1997), which is similar to the trend of the relative amount of CTV in the aphids. To understand the mechanisms of pathogen–vector interaction, a membrane feeding and immunofluorescent-localized lettuce infectious yellows virus assay was developed by Chen et al. (2011). The caged whiteflies were provided sequential access to an artificial liquid diet containing antibodies to the virion, followed by second antibodies conjugated with Alexa Fluor 488 for about 48 h in total after the acquisition feeding of virions (Chen et al., 2011). This assay does not need to create fluorescently labeled virions. On the basis of the remaining CTV amount in the aphids in this paper, the caged aphids were given access to an artificial liquid diet for no more than 12 h and even 3 h after the acquisition feeding of virions, while Chen et al.’s approach (2011) was used to analyze the correspondence between virion retention in the cibarium of insect vectors and aphid transmission of different CTV isolates. In addition, the CTV virions retained in aphids gradually decrease over time, which is a characteristic of semi-persistent transmission (Ng and Falk, 2006).
In summary, RPAP0 and EF-1α were the two most appropriate reference genes for RT-qPCR normalization in viruliferous aphids. This is the first study on CTV titer variation in aphids after different post-acquisition feeding times on healthy plants. The remaining CTV titer in the aphids showed a loss of about 80% for the highly transmissible isolate CT11A, while a loss of about 40% occurred for the poorly transmissible isolate CTLJ during the first 0.5 h of feeding time on healthy plants. To avoid CTV introduction and dispersal, aphid infestations developing on CTV-infected trees should be prevented.
Notes
Conflicts of Interest
No potential conflict of interest relevant to this article was reported.
Acknowledgments
This study was supported by the National Natural Science Foundation of China (Grant No. 31972237), the National Key R&D Program of China (Grant No. 2021YFD1400800), and the Fundamental Research Funds for the Central Universities (Grant No. XDJK2019B017).