



Orchid, which belong to the family Orchidaceae, is one of the most important ornamental crops in the world’s flower industry, because of their attractive flowers with diverse shapes and long vase life (Lawson et al., 1995; Li et al., 2019). In Korea, the annual production of orchids was 112 million pots, and the wholesale value was about 27.1 million US dollars (Ministry for Food, Agriculture, Forestry and Fisheries, Korea, 2021). Orchids have been reported to be infected with more than 50 viruses (Zettler et al., 1990). Among orchid viruses, cymbidium mosaic virus (CymMV) is the most prevalent, economically important in orchid production (Ajjikuttira et al., 2002; Ryu and Park, 1995).
CymMV is a member of the genus Potexvirus of the family Alphaflexiviridae and the virus has a positive-sense single-strand RNA genome of approximately 6.3 kb. The genome of CymMV consists of 5 open reading frames flanked by 5′ and 3′ non-coding regions plus 3′ poly(A) tail (Adams et al., 2004; Wong et al., 1994). Open reading frame (ORF) 1 encodes a 160-kDa replicase containing a methyltransferase domain in the N-terminal region, the RNA helicase domain and the RNA-dependent RNA polymerase domain in the C-terminal region. ORFs 2-4, referred to as the triple gene block, are involved in cell-to-cell movement and suppression of RNA silencing. ORF 5 encodes the 24-kDa viral coat protein (CP), which is indispensable for virus assembly and systemic spread in orchids (Verchot-Lubicz et al., 2007; Wong et al., 1994). CymMV frequently causes reductions in yield and quality of orchid flowers in Korea as well as in countries where orchids are cultivated (Sherpa et al., 2006; Wisler et al., 1987; Wong et al., 1994; Yoon et al., 2012; Zettler et al., 1990). Orchids infected with CymMV showed color-breaking or necrotic spots in flowers, and streak, yellowing or stripe mosaic in leaves (Wisler et al., 1987; Wong et al., 1994; Yoon et al., 2012; Zettler et al., 1990). Therefore, the detection of CymMV for the control of virus infection is important as it helps to reduce the economic losses in orchid production.
To date, some diagnostic methods have been developed to detect CymMV in orchid plants. These techniques include immunological assays (Eun and Wong, 1999; Park et al., 1998; Tanaka et al., 1997; Yoon et al., 2014), nucleic acid hybridization (Eun et al., 2002; Hu and Wong, 1998), reverse transcription (RT)-polymerase chain reaction (PCR) (Kim and Choi, 2015; Seoh et al., 1998; Yoon et al., 2011), and one-step RT loop-mediated isothermal amplification assay (LAMP) (Lee at al., 2011). However, all of these techniques have a few disadvantages, such as time-consuming, low-sensitivity, cross reactivity, laborious or requirement of expensive equipment. Most of these technologies are not applied directly to detect CymMV in the orchid farms, but have to be applied in the laboratory. For example, RT-LAMP is a reliable detection method. However, this technology requires four to six primers, and the primer design process is relatively complex. RT-LAMP is also limited to inapplicability for cloning, multiplex application and low specificity (Wong et al., 2018).
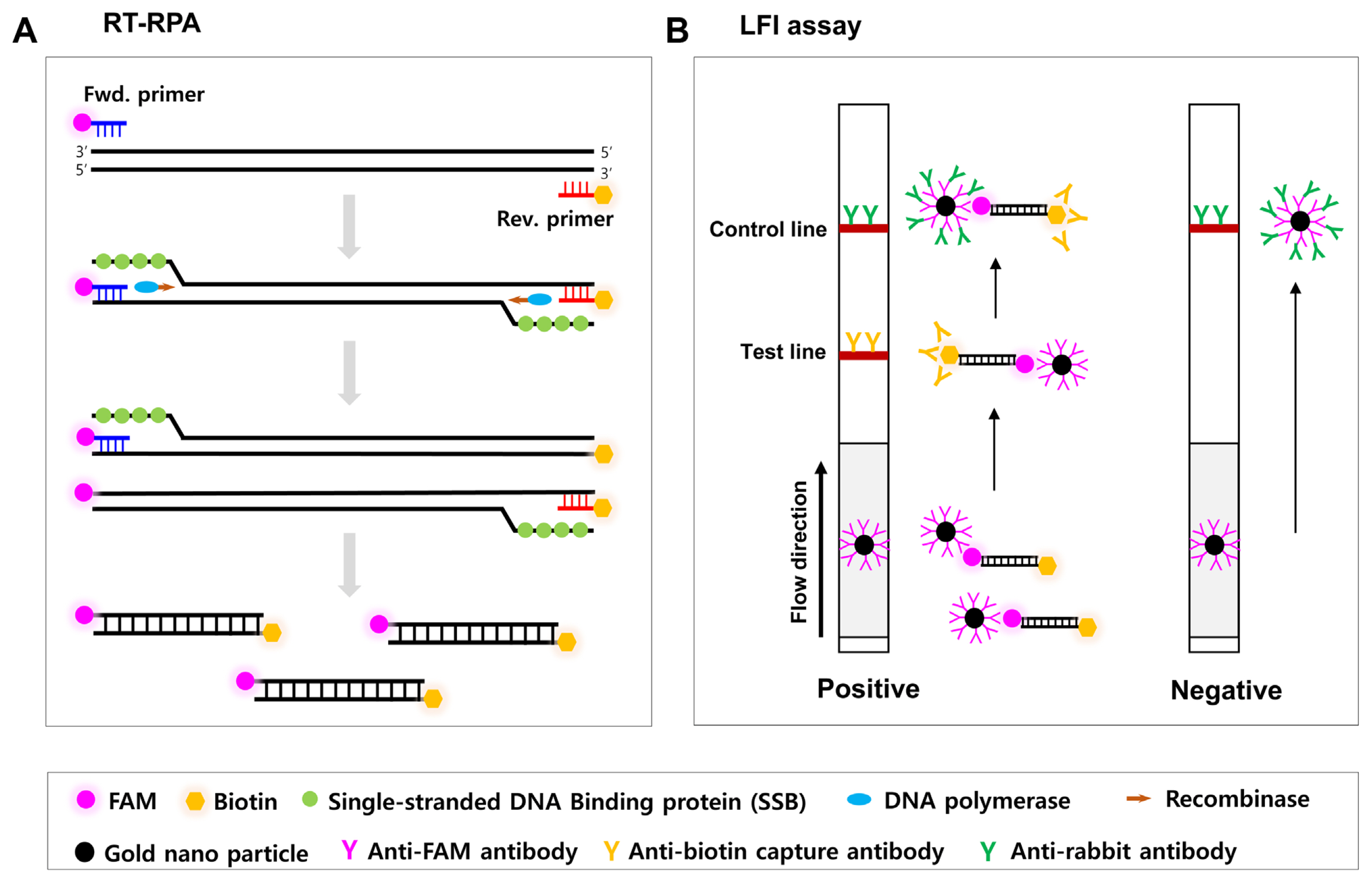
Recently, isothermal recombinase polymerase amplification (RPA) and RT-RPA assay have been shown to be a rapid, simple, sensitive and cost-effective approach to DNA and RNA detection in plant viruses and viroids (Babu et al., 2018; Ivanov et al., 2020; Kim et al., 2019, 2020; Kovalskaya and Hammond, 2022; Lodoño et al., 2016; Lu et al., 2018; Mekuria et al., 2014; Silva et al., 2014; Zhang et al., 2014). The RPA or RT-RPA can be generally carried out at low temperature (37-42°C) and the reaction is completed within 20 min depending on the starting copy number of the targeted template and the size of the amplicon (Li et al., 2019). In general, RPA products are analyzed using agarose gel electrophoresis, probe-based fluorescence or lateral flow immunoassay (LFI). Of the analyzing methods, RPA products combined with LFI (RPA-LFI) is able to avoid the time and equipment required for electrophoresis (Milenia Biotec, Giessen, Germany). To achieve the RPA-LFI technology, primers should be end-labeled with chemically modified reporter molecules, such as 5′-FAM or 5′-biotin. Subsequently, the resulting amplicon containing biotin and FAM can be shown using a nitrocellulose membrane coated with anti-biotin antibody and anti-FAM nanogold conjugates (Fig. 1). This study is aimed to develop a RT-PRA method for CymMV using specifically modified primers and LFI, and to validate for on-site CymMV detection in orchid samples from farms without any equipment.
Materials and Methods
Virus source
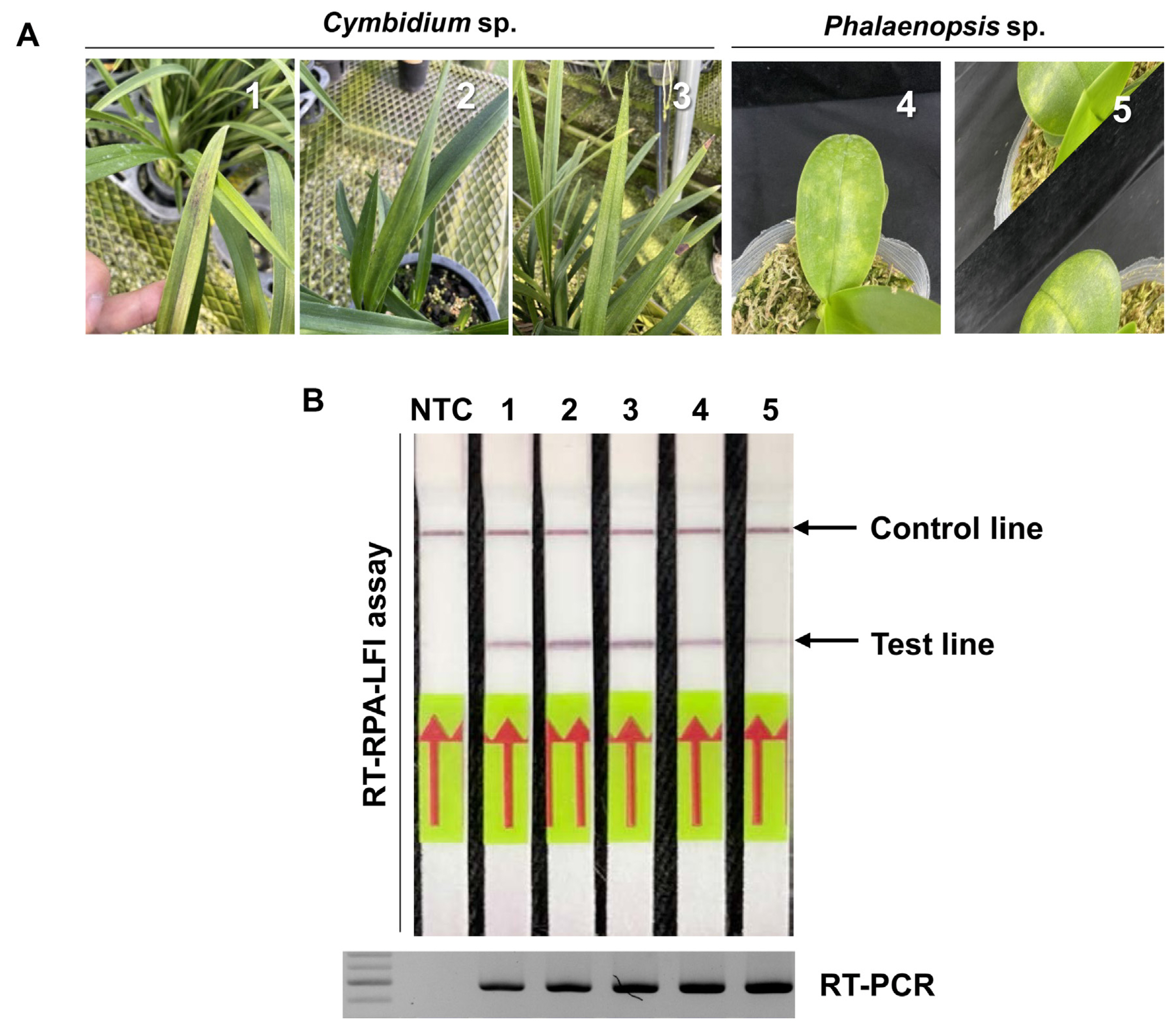
Cymbidium sp. and Phalaenopsis sp. which showed necrotic stripe or chlorotic patch symptoms were kindly obtained from National Institute of Horticultural and Herbal Science, Rural Development Administration, Korea in 2020. The plants were maintained in a growth chamber with a 16 h light/8 h dark cycle at 24°C.
Total RNA extraction and RT-PCR
Total RNA was extracted from symptomatic leaves of 8 Cymbidium sp. and 7 Phalaenopsis sp. using easy-BLUE total RNA extraction kit, according to manufacturer’s instructions (iNtRON Biotechnology Inc., Seongnam, Korea). Briefly, 100 mg of fresh leaf tissue was ground to a fine powder using liquid nitrogen, transferred to a sterile 1.5 ml tube followed by the addition of 1.0 ml of extraction buffer (100 mM Tris [pH 8.0], 50 mM EDTA, 500 mM NaCl, and 10 mM 2-mercaptoethanol). The tube was vortexed vigorously, and then incubated at room temperature for 5 min. Two hundred microliter of chloroform was added and vortexed vigorously, followed by incubation at room temperature for 3 min. The sample was centrifuged at 13,000 rpm for 15 min at 4°C. The upper aqueous phase was collected into a new tube and 0.5 ml of 100% isopropanol was added to the tube. The mixture then was incubated at room temperature for 10 min and centrifuged at 13,000 rpm for 10 min at 4°C. Total RNA was washed using 1.0 ml of 75% ethanol, and centrifuging the tube at 10,000 rpm for 5 min at 4°C. The RNA pellet was air-dried for 10 min. The RNA pellet was then resuspended in 50 μl of RNase-free water. RNA quality and concentration were determined using a NanoDrop spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA), and total RNA was stored at −70°C for use later.
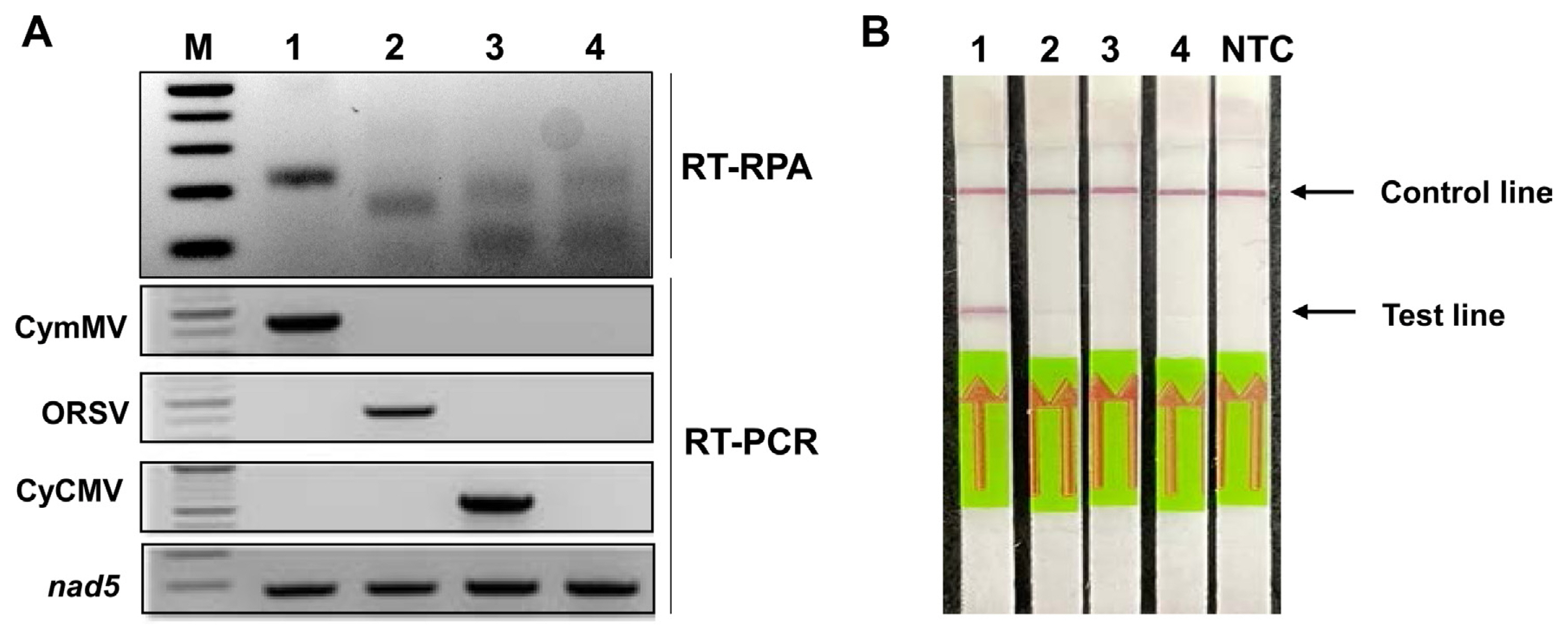
A single infection of CymMV in Cymbidium sp. and Phalaenopsis sp. was confirmed by RT-PCR with primers specific to CymMV, odontoglossum ringspot virus (ORSV), or cymbidium chlorotic mosaic virus (CyCMV), as describe previously (Chung et al., 2010; Yoon et al., 2018) (Supplementary Table 1). A healthy Cymbidium sp. was used as a negative control. Orchid mitochondrial nad5 mRNA was used as the internal control for RT-PCR assay (Lee and Chang, 2006) (Supplementary Table 1). RT-PCR was carried out using 1 μl of total RNA, 1 μl of each of 10 pmoles of forward and reverse primers, 13 μl of DEPC-treated water, and 5 μl of Topscript one-step RT-PCR mix (Enzynomics, Daejeon, Korea), with a total reaction volume of 20 μl. The thermo-cycling conditions were as follows: 30 min at 50°C for RT, 10 min at 95°C (1 cycle), 94°C for 30 s, 50°C for 30 s, and 72°C, for 40 s (40 cycles), and a final extension at 72°C for 5 min.
Primer design and RT-RPA-LFI assay
To design primers specific for RT-RPA-LFI assay, CP sequences and TGBp1 sequences of 13 CymMV isolates (AF016914, LC125633, AB198937, EF125180, AM055720, HQ681906, AY571289, U62963, AM055640, JQ860108, EU314803, MK816927, and KR185347) originating from some countries were downloaded from the GenBank database. Then, conserved regions of CP genes or TGBp1 genes were analyzed using multiple sequence alignments of MEGA10 (Kumar et al., 2018) (Supplementary Table 2). After designing a total of 10 primer sets targeting CP and TGB1, primer set showing the optimal RT-RPA result was selected among these primer sets, and biotin or FAM was attached to the end of the primers (CymMVT1-3sFAM-F and CymMVT1-3sBiotin-R) (Table 1, Supplementary Table 3). They were used for optimizing for RT-RPA-LFI. The RT-RPA-LFI carried out with those primer set according to the manufacturer’s instructions of TwistAmp Basic kit (TwistDX, Ltd., Cambridge, UK). The RT-RPA-LFI assay for CymMV detection in orchids was performed in a 50 μl reaction volume containing 1 μl of total RNA isolated from each CymMV-infected orchid tissue, 29.5 μl rehydration buffer, 2.4 μl of 2 pmoles forward primer, 2.4 μl of 2 pmoles reverse primer, 1.0 μl of RevertAid reverse transcriptase (Thermo Fisher Scientific), 2.5 μl of 280 mM magnesium acetate, and 11.2 μl DEPC-treated water. After a brief spin, the reaction was incubated for 30 min at 38°C in heat block with a closed lid. For lateral flow assay, with reference to the Milenia GenLine HybriDetect 1 (Milenia Biotec), 10 μl of RT-RPA products were mixed with 100 μl of assay buffer (HybriDetect assay buffer, Milenia Biotec) in a new reaction tube. Then the LFI strip was immersed into the mixture and incubated for 5 min at room temperature. A band with one visible line in the control line was considered negative, and a band with two visible lines in both the control line and the test line was determined as positive (Fig. 1B, Supplementary Fig. 1).
Results
Detection specificity of RT-RPA-LFI assay for the detection of CymMV
The detection specificity of the primers was verified in a single CymMV, ORSV, or CyCMV-infected orchid sample by RT-RPA assay (Fig. 2A). The RT-PCR was performed to verify the viral presence on the orchids (Fig. 2A). The RPA-LFI assay successfully detected CymMV in each orchid using primers specific to the partial TGBp1 gene of CymMV, and did not detect CymMV in ORSV-infected, CyCMV-infected and a healthy orchid plant, similar to the results of RT-RPA on agarose gel electrophoresis (Fig. 2B). These results suggest that the specificity of RT-RPA for CymMV detection is equivalent to that of conventional RT-PCR analysis and has no cross-reactivity in ORSV and CyCMV at the test band of the RT-RPA-LFI assay.
Optimization of the RT-RPA-LFI assay
The conditions of the RPA reaction were optimized based on the reaction temperature and time. The assay was used with a selected primer set (CymMVT1-3s-F and CymMVT1-3s-R) and visualized on the 2% agarose gel after 15 min of RT-RPA incubation time at a temperature of 35-42°C (Supplementary Fig. 2). The RT-RPA-LFI assay was performed on CymMV-infected Cymbidium leaves at 1, 5, 10, 20, and 30 min, respectively. The assay was visualized with a red band at the test line of the immunostrip from 10 min onwards and remained until 30 min at a temperature of 39°C. The strips with no-template control (NTC) were not visualized any line in the same range of time (Fig. 3). Therefore, it was determined that the optimal reaction conditions for the detection of CymMV RT-RPA-LFI assay were 39°C for 15 min.
Application of the RT-RPA assay for CymMV detection
To survey the CymMV infection, a total of five samples were randomly selected among 8 Cymbidium sp. and 7 Phalaenopsis sp. plants and subjected to RT-RPA-LFI assay (Fig. 4A). All leaves have confirmed the infection of CymMV by RT-RPA-LFI assay without a false-positive band in the test line and consistent with the RT-PCR results (Fig. 4B). These results indicated that the RT-RPA-LFI assay could be used reliably for CymMV detection in field-collected samples.
Discussion
In the present study, we successfully developed a RT-RPA-LFI assay to detect CymMV from orchid plants, showing a simple, rapid, sensitive, and accurate method. The RT-RPA-LFI assay for CymMV detection was not showed cross-reactivity with three orchid-infecting viruses (Fig. 2). On-site detection of CymMV from orchid leaves in farms proved applicability of the RT-RPA-LFI assay and the determination for the presence of CymMV in farms can be verified from 10 minutes after the reaction, compared with 150 min for the RT-PCR assay. These features allow wide use both in the laboratory and in the field.
To date, there are a number of reports on the development of RPA-based assays for plant viruses (Babu et al., 2018; Babujee et al., 2019; Kim et al., 2019, 2020, 2022, 2016; Lu et al., 2018; Mekuria et al., 2014; Silva et al., 2014; Zhang et al., 2014) and viroids (Hammond and Zhang, 2016; Ivanov et al., 2020; Kovalskaya and Hammond, 2022; Lee et al., 2020). The RT-RPA-LFI assays for detection of plant viruses and viroids were performed in accordance with the TwistDx kit and lateral flow strips were utilized as an end-point readout method (Kim et al., 2022; Kovalskaya and Hammond, 2022). Most RPA assays were developed with a TwistAmp nfo kit which is required nfo probe as well as a specific primer set (Babu et al., 2018; Ivanov et al., 2020; Kim et al., 2022). Here, we validated RPA reaction for the detection of CymMV with a TwistAmp basic kit without a probe. To decrease the excessive amplification in RPA reaction, half the concentration (4.8 pmole) of primer was used and RPA generates a limited amount of the target. The low quantity of amplicons was not problematic as a non-specific reaction in the NTC reaction. To our knowledge, this is the first report of the development of RT-RPA-LFI assay to detect CymMV RNA in orchid plants using TwitAmp basic kit, combined with the LFI method.
Detection methods for CymMV include generally double-sandwich enzyme-linked immunosorbent assay (DAS-ELISA) (Park et al., 1998; Sherpa et al., 2006), nucleic acid hybridization (Eun et al., 2002; Hu and Wong, 1998), RT-PCR (Chung et al., 2010; Park et al., 1998; Seoh et al., 1998; Yoon et al., 2012), multiplex RT-PCR (Ali et al., 2014; Kim and Choi, 2015; Lee and Chang, 2006), RT-LAMP (Lee et al., 2011), and real-time RT-PCR (Eun et al., 2000). Most of these assays require expensive equipment, purification of nucleic acids, trained persons and analytic tools of amplified products by gel electrophoresis or specialized equipment. Furthermore, commercial DAS-ELISA kits and lateral flow immunoassay kits for CymMV and ORSV are available from a few companies. The latter kits have been known for on-site diagnosis of CymMV and ORSV, similar to the RT-RPA-LFI assay for CymMV detection. Nonetheless, one of disadvantages of the RT-RPA-LFI assay is the cost. The price of RT-RPA reagents per sample is higher than that of RT-PCR analysis. There have been some reports for cost cut-off per sample in lateral flow strip detection, such as in the use of inexpensive paper and glass fiber (Lobato and O’Sullivan, 2018). Development of the simple sensitive, rapid and accurate RT-RPA-LFA assay helps early diagnosis and subsequent prevention of CymMV in orchid production in Korea.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement
Supplement Print
Print






