


Pear scab, caused by Venturia nashicola (Tanaka and Yamamoto, 1964), is an economically important disease in Korea (Cho et al., 1985). The pathogen is different from the European pear scab fungus (Venturia pirina). V. nashicola and V. pirina are differentiated by their morphological and pathological characteristics, and each species operates in its own Venturia-Pyrus pathosystem: V. nashicola co-habits with the commercial Asian pear species (P. bretschneideri, P. ussuriensis, and P. pyrifolia), while V. pirina has followed the dissemination of the European pear (P. communis) to many parts of the world (Ishii and Yanase, 2000). V. nashicola infects leaves, fruits, and young shoots, resulting in significant annual yield loss of pear production in East Asia (Abe et al., 2008; Ishii and Yanase, 2000; Park et al., 2000; Tanaka and Yamamoto, 1964). Pear scab can damage stems, petioles, and apical shoots at the flowering stages through maturity. The disease causes deformity and failure of uniform enlargement of fruit. Also, leaves and stem infection by V. nashicola reduced yield and quality in the following year. The pathogen can overwinter as conidia in buds on pear trees as dormant mycelia, in dead leaves on the ground as immature pseudothecia, and on the surface of twigs or diseased leaves (Holb et al., 2004; Ishii and Yanase, 2000; Lian et al., 2007). The main form of primary inoculum in most pear growing areas during the spring is regarded as the conidia produced from the overwintered dormant mycelia in the buds and on the surface of previously infected twigs or dead leaves (Lian et al., 2007). Because the pear scab is not readily controlled by fungicides, effective control in the field can be achieved by growing resistant cultivars or by removing overwintering infected buds, twigs, and diseased leaves as often as needed to reduce the disease pressure (Bell and van der Zwet, 1998). However, pear scab in Korea is currently controlled by scheduled application of fungicides, as there has been limited research on biology and resistance to pear scab. Actually, few commercial pear cultivars carry moderate levels of resistance to V. nashicola. Furthermore, resistant cultivars providing potential for resistance breeding usually display low fruit quality and are not suitable for commercial use (Postman et al., 2005; Won et al., 2014). In contrast, a few Asian pear cultivars, which are generally susceptible to V. nashicola, are immune to infection by V. pirina because of non-hosts for Western species. Genetic resistance can be the most effective mean to maintain fewer control input and lower production cost when pear scab occurs (Postman et al., 2005; Won et al., 2014). Only a few genes involved in pear scab resistance can be found in Asian pear (Brewer et al., 2009). In Asian pear, scab resistance to mixed isolate inoculum of V. nashicola was considered to be controlled by a single dominant gene (Abe and Kotobuki, 1998a). However, the same authors observed necrotic reactions characterized by symptoms without sporulation on some Asian pear cultivars and suggested that necrotic reaction to V. nashicola may be controlled by multiple genes of small effect (Abe and Kotobuki, 1998b). In the present study, we compared the traditional dropping inoculum method with an alternative inoculation method using mycelial plug to suggest a fast and reliable standard for determining disease-resistant plants. Resistance was evaluated by comparing susceptible and resistant cultivars inoculated by both methods.
To evaluate pear response to V. nashicola, we generally collected scab lesions from diseased leaves of susceptible pear cultivar grown in a commercial orchard and washed with distilled water to make a conidial suspension. Then, the final suspensions were adjusted to a concentration of more than 1 × 105 conidia/ml for the infection experiment. In addition, diseased leaves were collected from other orchards in the same location to get enough conidia. Hence, the final resulting suspension was a mixed inoculum. This method does not allow for reproducible observations of pear responses to V. nashicola because of limitation of the same kind of inoculum collection.
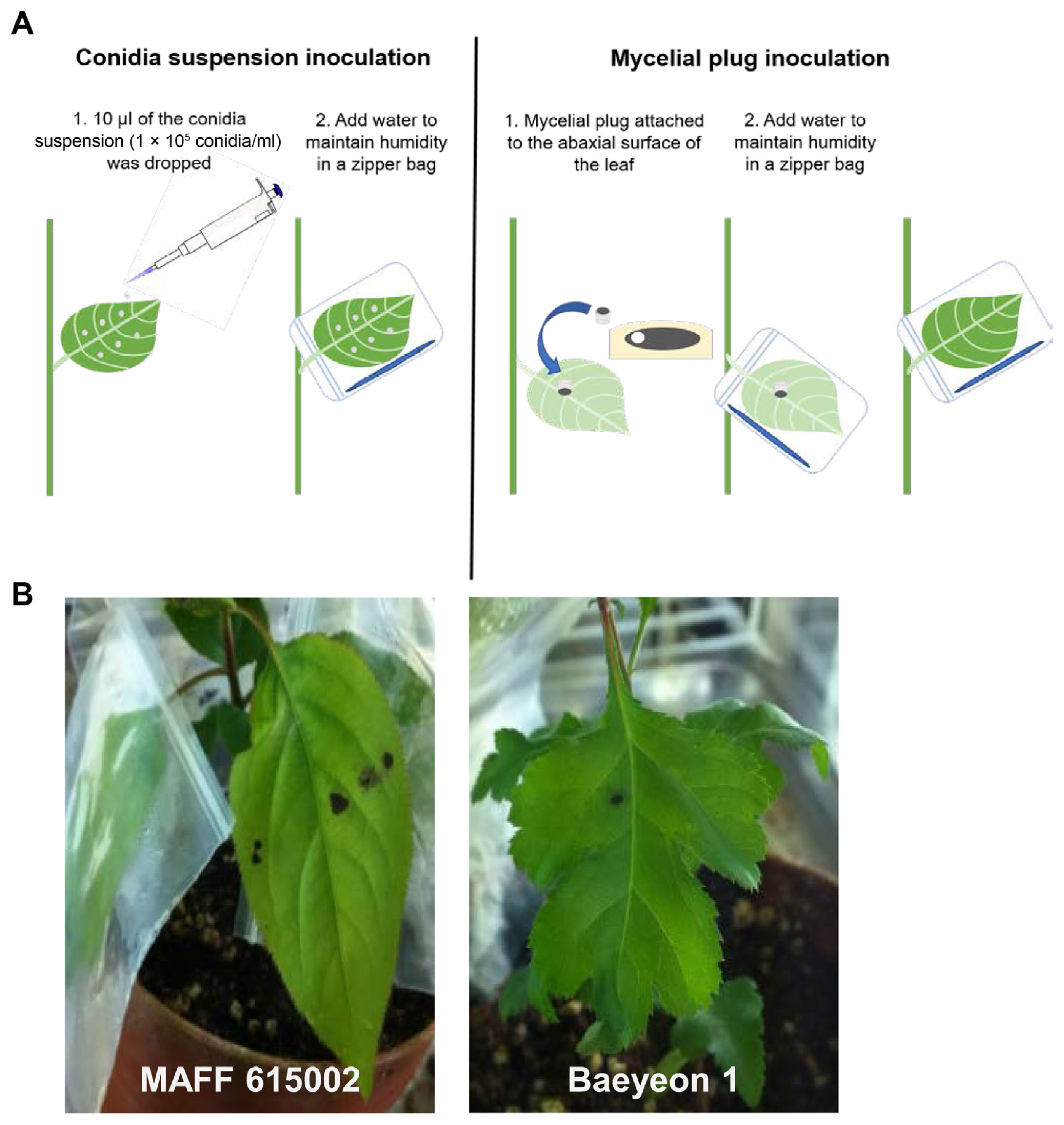
Single spore isolation is essential for studies of plant response as well as fungi itself. To isolate single conidia of V. nashicola, leaf, leaflet, fruits, and twigs with pear scab symptoms were collected on pear trees from the field of the department of Pear Research Institute (National Institute of Horticultural and Herbal Science) in Naju, Korea (Fig. 1A and B). Pear twigs containing a lot of conidia were put on stage and fixed with a stage clip to the light microscope without a light condenser. Hand-made glass needles were jointed on a holed cork itself fitted into a condenser lens socket instead of a condenser. Conidia of V. nashicola were picked up at the tips of glass needles by using the condenser knob and dragged onto 2% water agar plate. Hundred isolates were cultivated until the emerging colonies were visible at 22oC in the dark. Colonies of V. nashicola (individual colony designated as Baeyeon 1-24) were then transferred and cultivated onto potato dextrose agar (PDA) for at least 2 months (Fig. 1D). The obtained pure cultures, Baeyeon 1-24 were used for studies of conidia formation and pathogenicity assay.
Two pear cultivars, Pyrus pyrifolia cv. Hwasan with known disease susceptibility and Pyrus hybrids (P. pyrifolia × P. communis) cv. Greensis as resistance control, were used for evaluating the response to pear scab caused by V. nashicola in spring, and then maintained in a greenhouse. One-year-old grafted pear plants were used for inoculation. These cultivars of pear were tested for disease response by dropping conidia inoculation or direct fungal plug inoculation methods. Isolates of V. nashicola obtained from single spore isolation or MAFF 615002 from the department of Pear Research Institute were grown on PDA medium in Petri plates at 22°C for 2 months. After 60 days cultivation, colonies of V. nashicola were cut into tiny pieces using a sterile razor blade or a sharpened inoculation needle with 10 ml of sterile tap water. One point five-1 ml of fungal suspension was applied on the surface of 1% Cheongah (Jangmyeog Food Co., Seoul, Korea) agar media (Choi et al., 2017) to induce conidia production of V. nashicola and the fungal suspension was gently spread using sterile glass-rod shaped like a hockey stick to get almost same age of colonies. After drying the plates in a clean bench, isolates of V. nashicola were cultivated for 45 days at 22°C in the dark until producing conidia (Eguchi and Yamagishi, 2008). Mycelial growth was slower but covered almost all the area of the Petri dish containing 1% Chenongah medium 45 days after cultivation. First, we observed conidia of V. nashicola MAFF 615002 30 days after cultivation on Cheongah medium, but only 3 isolates out of 24 single isolated colonies were found on the same media. The number of conidia produced was about 1 × 105 conidia/plate 30 days after the cultivation of V. nashicola MAFF 615002, while newly isolated V. nashicola produced less than 4.5 × 103 conidia/plate. The isolate Baeyeon-1, produced the most conidia among the three isolates that were employed for the next disease assessment.
Conidia of V. nashicola were harvested from 30-day-old 1% Cheongah medium, washed with distilled water, and suspended in 0.01% Tween 20. The conidia concentration was adjusted to 105 conidia/ml. Ten microliters of conidia suspension were placed on the upper side of each leaves. After air drying, the inoculated leaves, were sealed in a plastic zipper bag in order to maintain the moisture conditions favorable for penetration of fungus into the leaf tissue (Fig. 2A). The inoculated plants were placed in a growth chamber and maintained at 25°C. All the leaves were consistently infected and developed symptoms at the inoculation point after infection with MAFF 615002 or the newly isolated strains. Small dark brownish lesions were observed on the leaves of cv. Hwasan 14 days after inoculation (DAI). Dark lesion of 5 mm diameter was observed in the all plants inoculated with tested isolates, however, there were no extending symptom 3-4 weeks after inoculation (Fig. 2B). The dropping of conidia suspension to the leaves has the following disadvantages: (1) it is not reliable because conidia suspensions may not stay on the leaf, (2) it is time-consuming to make conidia on special media, (3) it required at least 2 weeks for determining pathogenicity. Conidia dropping method is not suitable for accurate disease assay especially for estimating disease responses against pear scab. An effective inoculation method is a critical component of an accurate disease assay for quantifying levels of pear scab resistance among pear cultivars. The type of inoculum used in an inoculation method is a major determinant of infection efficiency. Environmental and pathogenic factors, e.g., changes of humidity, the vitality of inoculum, types of inoculum, and so on, could lead to additional variation in the infection process and subsequent disease development (Li et al., 2003). Therefore, using an active growing source of V. nashicola inoculum is important. So, we changed the inoculum to mycelium plug instead of conidia of V. nashicola to have easy and effective inoculation method. Pear cv. Hwasan at the eight-leaf stage were inoculated with V. nashicola by placing round mycelial plug (1 cm of diameter) on the abaxial surface of the leaves to enhance mycelia invasion through stomata (Fig. 2A). The inoculated leaf was sealed in a zipper bag. The main purpose of developing this technique was to enable screening single isolates of V. nashicola in pear cultivar at an early stage. As an additional control, we selected “Grensis”, another pear cv. is already known to show field resistance to pear scab disease.
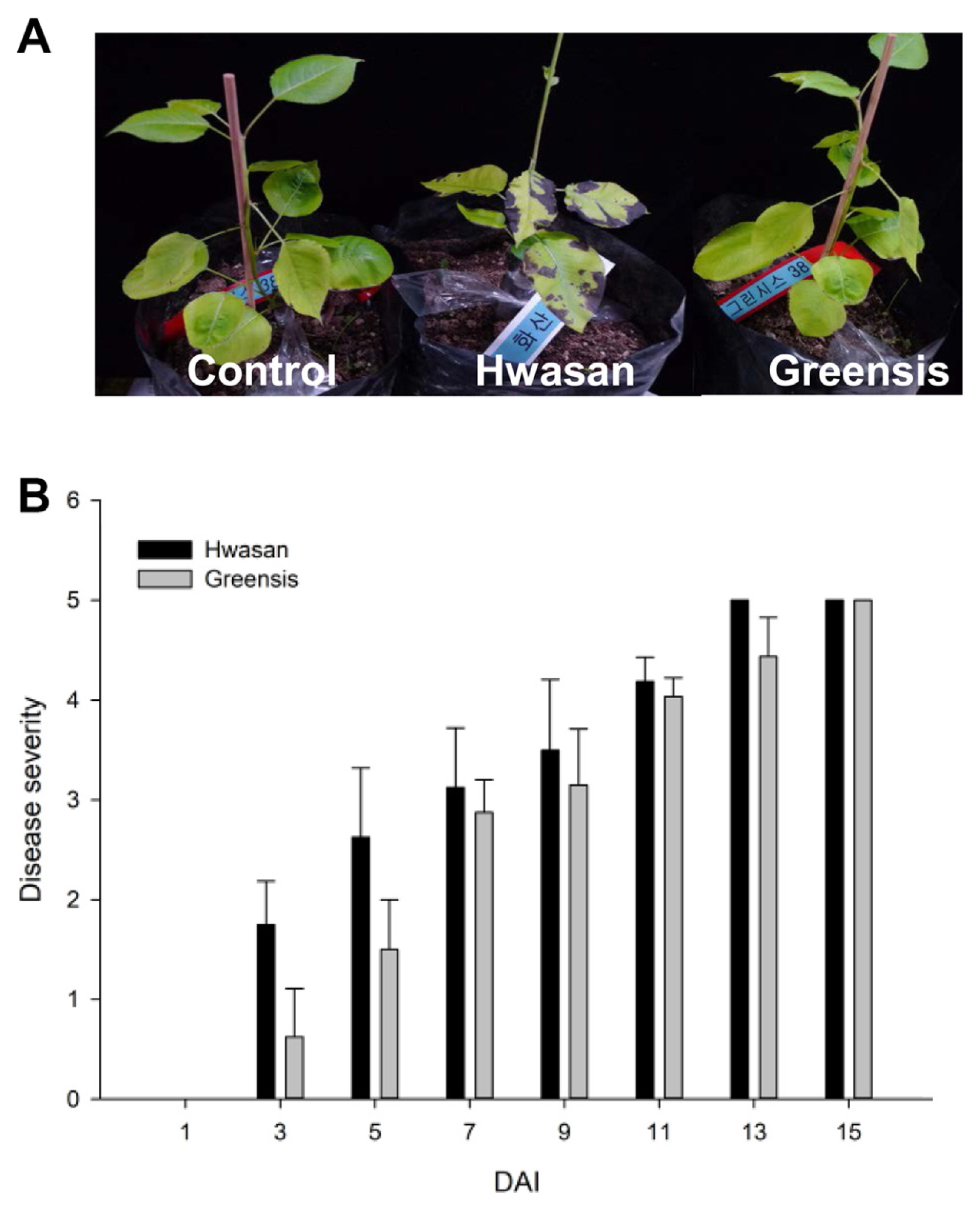
When inoculated with mycelial plugs of different isolates of V. nashicola, susceptible pear cv. Hwasan had a dark brownish leaf-discoloration rapidly extending from the inoculation site preceding defoliation. Leaf lesions rapidly expanded to form round or irregularly shaped lesions with all tested isolates of V. nashicola. The severity of infection was not affected by the position of the leaves; all leaves developed symptom to the same extent. To standardize disease symptoms on the leaves, disease severity was rated daily after inoculation based on a 0 to 5 scale, where 0 = no visible disease symptoms; 1 = brownish lesions beginning to appear on the leaves; 2 = approximately 21-40% of lesion on the leaf; 3 = approximately 41-60% of symptom on the leaf; 4 = approximately 61-80% of symptom on the leaf; 5 = leaf defoliated, more than 81% of symptom on the leaf or dead (Fig. 3). Pear germplasm at the U.S. Department of Agriculture (USDA) Genebank has been evaluated for scab resistance by both controlled inoculation and observation of natural symptoms based on 1 to 9 scale rating (U.S. Department of Agriculture, 2010). However, it was challenging to resolve the range of symptoms at the juvenile stage of the pear plant because of the rapid developing blight symptoms in our experiments.
Expression of resistance in pear plants to V. nashicola was affected by the inoculation method and the incubation period. With traditional inoculation by dropping, differences in the symptom severity were observed between the two cultivars at the eight-leaf stage of pear plants. Scab symptoms were slightly induced on the susceptible cv. Hwasan at 14 DAI, while symptoms were not visible on the resistant cv. Greensis at this stage. Lesions induced by dropping inoculation on the susceptible cv. Hwasan were developed around the dropped inoculum area at 14 DAI. Mean lesion diameter on the susceptible cv. Hwasan increased from 0.5 cm to 1.0 cm by 28 DAI with typical stellated symptoms, while the lesions started to develop at 22 DAI in the resistant cv. Greensis and lesion development was much slower than susceptible cv. Hwasan (data not shown).
Direct attachment of the mycelial plug of V. nashicola resulted in scab symptoms on both susceptible cv Hwasan and resistant cv. Greensis. But the incubation period of V. nashicola in the resistant was longer than that in the susceptible. Disease severity on Hwasan and Greensis differed at the juvenile stage of pear. Disease severity was rated according to the portion of leaf blighted (Fig. 3), and lesion development in the susceptible cv. Hwasan was progressively more severe than in the resistant cv. Greensis within 3-9 days (Fig. 4A and B). Resistant cv. Greensis displayed a mean disease rates of 0.6, 1.5, 2.8, and 3.2 at 3, 5, 7, and 9 DAI, respectively. While susceptible cv. Hwasan displayed a mean disease rates of 1.7, 2.6, 3.2, and 3.5 at 3, 5, 7, and 9 DAI, respectively. However, the interval of disease severity between susceptible and resistant plants was decreased over time after inoculation, and the disease severity peaked at 15 DAI in both pear cultivars. This suggests that attachment of mycelial plug would provide a fast standardized method for determining resistant plants during early stages of growth, but longer trials would not guarantee the accuracy for selecting disease-resistant plants. Taken together, the differences between the susceptible and resistant responses to V. nashicola were more quantitative than qualitative, because some symptoms developed slowly on resistant pears. We also found that resistance of pear scab could be overcome by the prolonged exposure to the fungus.
Disease-resistant cultivars are the most economically and environmentally beneficial way of reducing loss to pear scab. Cultural control methods are insufficient and fungicides may not be economically or environmentally sustainable. A concerted effort is currently in progress in pear-growing countries to identify genes responsible for pear scab resistance and to study the key genes and underlying mechanisms of the disease resistance (Postman et al., 2010). To achieve these goals, uniform, easy and effective infection and accurate evaluation methods are required for genetic, molecular, and biochemical analysis and for measuring quantitative differences in pear scab resistance among pear breeding lines, mutants, and transgenic plants. Most recently, a useful method has been reported to produce conidia on special media (Choi et al., 2017). In this study, we developed an improved method for consistent and reproducible inoculation of pear scab disease and for more accurate measurement of disease severity.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







