



Tuber mustard (Brassica juncea Coss. var. tumida Tsen et Lee) is an economically and agriculturally important crop belonging to the Cruciferae family which is cultivated in South-East Asia (Qi et al., 2008). Due to special flavor and nutritional value, the succulent stems of tuber mustard are used in pickled products with a high popularity in China. White mold is one of the most important diseases of tuber mustard production in Zhejiang province, China. The causal pathogen, Sclerotinia sclerotiorum (Lib.) de Bary, is a destructive soilborne fungus which can infect a wide range of plant species in the world (Boland and Hall, 1994).
To date, no resistant cultivar has been developed against white mold of tuber mustard and application of different fungicides is a major method used by local growers against the disease. A large number of researches are annually conducted to manage sclerotinia diseases on different plants. Previous studies have shown that biological control (Ojaghian, 2010, 2011), Brassica biofumigation (Ojaghian et al., 2012), UV-C radiation (Ojaghian et al., 2017) and application of resistance inducers (Ojaghian et al., 2013) have potential to reduced severity or incidence of S. sclerotiorum on different crops. However, the majority of local growers prefer applying chemical fungicides to natural substances or biocontrol agents which has caused significant concerns over environmental conditions.
Chitinases play a major role in defense reaction to fungal phytopathogens because chitin is a main constituent of fungal mycelial walls (Neuhaus, 1999). Chitinases are chitin-degrading enzymes with low-molecular weight which hydrolyze the ╬▓,1-4 linkages of chitin. Baranski et al. (2008) showed that the gene chit36 from Trichoderma harzianum enhances resistance of transformed carrot (Daucus carota L.) to Alternaria radicina and Botrytis cinerea. Furthermore, introducing chimeric chit42 from T. harzianum to carrot markedly reduced the pathogenicity of S. sclerotiorum (Ojaghian et al., 2018).
The current study was conducted to determine the effect of chit42 expression in tuber mustard plants against white mold.
The binary plasmid pGisPEC1 (Fig. 1) used in this study was developed by Ojaghian et al. (2018). Disinfection of the seeds of tuber mustard was done using ethanol (15%) and then 0.2% (v/v) hypochlorite sodium for 2 min. Tuber mustard seeds (cv. SCZX) were provided by College of Horticulture, Zhejiang University. The seeds were rinsed three times with sterile distilled water (SDW). According to Murashige and Skoog (1962), the seeds of tuber mustard plants were transferred on Murashige and Skoog (MS) medium. Formed cotyledonary petioles detached from germinated plants were placed on solid MS treated with 5 mg/l benzylaminopurine for pre-culture. After 5 days, the explants were used for transformation. Agrobacterium tumefaciens (HANG287) used in plasmid construction was grown in lysogeny broth including 50 mg/l kanamycin at 26┬░C to mid-log phase. Then it was transferred and grew in a fresh medium to OD600=0.4. After centrifuging at 5,000 ├Śg for 5 min, the collected bacteria were re-suspended in MS medium as explained by Ojaghian et al. (2018). The explants submerged in Agrobacterium-harboring strain for seven min were dried out on the sterile filter papers and placed on solid MS including 5 mg/l benzylaminopurine in a dark incubator at 24┬░C for 5 days. The explants were washed with SDW including 350 mg/l cephatoxime and then placed on the solid MS amended with 5 mg/l benzylaminopurine, 300 mg/l cephatoxime and 18 mg/l kanamycin. After shoot formation was initiated, the explants were transferred to solid MS and the regenerated shoots were detached from the explants and transferred to MS solid medium including 2.5 mg/l 3-indolebutyric acid and 300 mg/l cephatoxime to become conditioned for root formation. The media described above contained 12 g/l agar and 5% (w/v) sucrose. The explants were kept at 28 ┬▒ 1┬░C and 18 h photoperiod. The control explants were treated with pure SDW. The produced seedlings at 2-leaf phase were cultivated in the mineral soil in a greenhouse at 26┬░C for 75 days. In order to determine the presence of nptII and chit42 genes in tuber mustard plants, DNA was isolated from leaves of the transformed plants and controls as explained by Deng et al. (1995). Polymerase chain reaction (PCR) method was conducted using the specific primers and thermocycler adjustment described by Ojaghian et al. (2018). The PCR products were electrophoresed on an ethidium stained agarose gels. This test was done twice with two replicates for each sample. Moreover, southern blot test was employed to confirm introduction of the transgene to tuber mustard. Genomic DNA isolated from the leaf tissues of transgenic and control plants were digested with HindIII according to Ojaghian et al. (2018). After electrophoresis on agarose gels, the digested DNA was blotted to a nylon membrane (Amersham Hybond NTM+, GE Healthcare, Little Chalfont, UK). The probe was produced by PCR with the primers for chit42 and purification using a QIAEX II gel isolation kit (Qiagen, Hilden, Germany) as described by Ojaghian et al. (2018). In addition, DIG DNA Probe Kit (Roche, Shanghai, China) was used to label the PCR product. Hybridization was done at 42┬░C and the detection was done by DIG-High Prime DNA Labelling and Detection Starter Kit II (Roche) as described by Ojaghian et al. (2018). The southern blot test was repeated twice.
Specific mRNA of the transgene was checked using reverse transcriptase (RT)-PCR. Total RNA was extracted from the leaf tissues of four 75-day-old lines including TMB4, TMB7, TMB12, and TMB18 and control plants using Trizol according to the producerŌĆÖs manual (Sangon Biotech, Shanghai, China). First strand cDNA was generated using the oligo(dT)18 primer by using the ŌĆśŌĆśfirst strand cDNA synthesis kitŌĆÖŌĆÖ (Thermo Fisher Scientific, Waltham, MA, USA), which contained M-MuLV reverse transcriptase. PCR amplification was conducted using the first strand cDNA as template and chit42 specific primers under the same conditions described by Ojaghian et al. (2018). The analysis of RT-PCR was repeated twice.
In order to evaluate chitinase activity in the transgenic plants, protein was isolated from leaf tissues of the four lines including TMB4, TMB7, TMB12, and TMB18 according to Roberti et al. (2008). The activities of different kinds of chitinases were assessed in the lines TMB4, TMB7, TMB12, and TMB18 with three replications as described by Ojaghian et al. (2017). Finally, the effect of introducing chit42 on white mold was determined in the lines TMB4, TMB7, TMB12, and TMB18. Three isolates of S. sclerotiorum including Ss5, Sp2, and Sk1 were sampled from sclerotinia infected mustard fields in Hangzhou, China. High aggressiveness of the isolates was confirmed in a prior study using the straw inoculation method (Ojaghian, 2010). RAPD (random amplification of polymorphic DNA) test showed that these isolates were genetically different (data not published). The fungal isolates were cultured on potato dextrose agar (39 g/l; Merk, Darmstadt, Germany) and kept at 4┬░C until used.
A 5 mm mycelial plug was placed at the center of the plant leaf with the mycelium towards leaf surface. The leaves were placed in 10 cm plates on two sterile filter papers soaked with SDW. The filter papers were used to provide necessary humidity in the plates. The plates were kept in a dark incubator at 21┬░C. The infection (water soaked) radius on leaves was determined seven days after inoculation. This experiment was repeated three times for the three pathogen isolates. Using SAS software (SAS 8.2, 1999-2001, SAS Institute Inc., Cary, NC, USA) the effects of introducing chit42 against white mold caused by three pathogen isolates were assessed by ANOVA in a completely randomized design test. Hartley's Fmax test was used to prove the homogeneity of variance before running the statistics. Means of treatments were separated using FisherŌĆÖs least significant difference test.
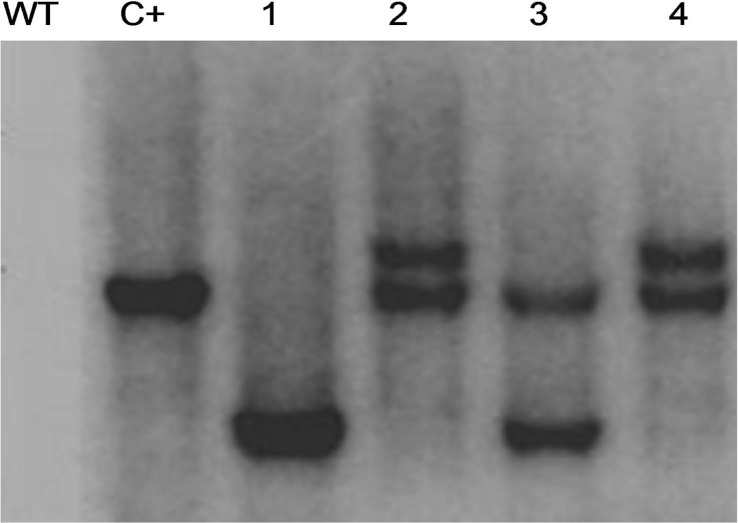
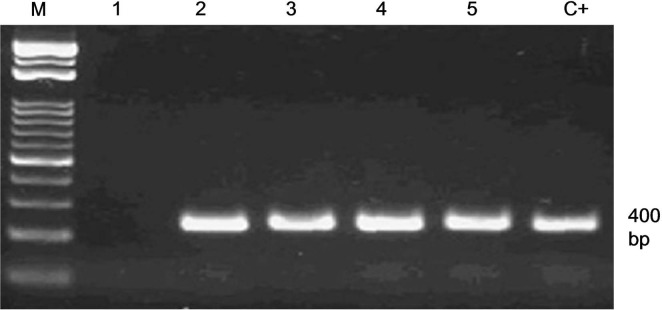
From 387 co-cultured explants 116 plantlets (29.9%) regenerated on MS medium, and 56% (65 out of 116) of the plantlets maintained green on the MS containing kanamycin. A number of 61 shoots showed successful rooting which transferred to soil. The PCR analysis showed that the gene chit42 was amplified in 27 lines which was equal to 6.9% (27 out of 387). Moreover, the gene chit42 was not amplified in the control (Fig. 2). In order to assure no Agrobacterium contamination possibly escaped the selection, genomic DNA from transgenic plants with virG primers was PCR amplified which resulted in no band after electrophoresis (data not shown). The results of southern blot showed integration of the transgenes in four randomlyselected PCR-positive plants (TMB4, TMB7, TMB12, and TMB18). As shown in Fig. 3, one, two, two, and two chit42-related bands were observed in the transformed lines TMB4, TMB7, TMB12, and TMB18, respectively. In addition, no hybridization sign was detected in the control plants. Following RT-PCR analysis, transgene mRNA expression was found in all four selected lines harboring chit42 gene. However, no accumulation of chit42-encoding mRNA was found in untransformed control plants (Fig. 4).
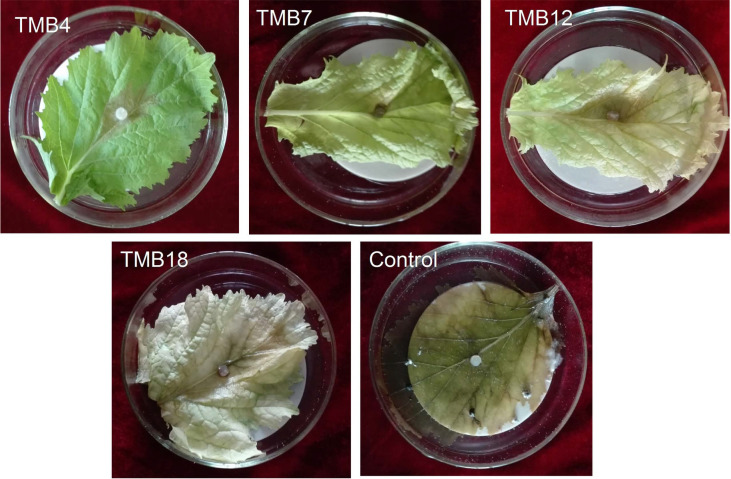
The results of enzymatic examinations showed a considerable increase in the activity of endochitinase in the four tested lines at a statistically similar level (Fig. 5). The level of endochitinase from 0.02 in the control increased to 0.052, 0.05, 0.051, and 0.053 U/mg protein in the lines TMB4, TMB7, TMB12, and TMB18, respectively. However, no significant change was observed in the activity of ╬▓-Nacetyl hexosaminidase and chitin 1,4-╬▓-chitobiosidase in the transformed plants compared with the control (Fig. 5). Furthermore, infection radius caused by three isolates of S. sclerotiorum markedly reduced in the four tested lines with statistically similar performance (Table 1, Fig. 6).
This study showed that the severity of white mold caused by S. sclerotiorum can be markedly reduced in tuber mustard plants due to introducing chit42. According to previous researches, the plants transformed by chitinase genes originating from plants (Punja and Raharjo, 1996) or Trichoderma spp. (Baranski et al., 2008; Ojaghian et al., 2018) show high level of resistance against different phytopathogens. This is the first study showing that the severity of white rot caused by S. sclerotiorum can be markedly reduced in tuber mustard plants due to introducing chit42. However, upregulation of the chit42 did not significantly reduce the pathogenicity of S. sclerotiorum in potato plants (cv. Agria) (data not published). Liu et al. (2011) introduced a chitinase PjChi-1 gene to Brassica napus which significantly enhanced the resistance toward Plutella xylostella and S. sclerotiorum.
Although chitinase activity showed a high range from 0.6 to 6 U/╬╝g in oilseed rape plants transformed by chit42 (Zarinpanjeh et al., 2016), chitinase activity increased in all tested transgenic tuber mustard plants at a statistically similar level in the present study. In addition, transformation efficiency was 6.9% that was much more than that tested by Gentile et al. (2007). However, it was similar to the transformation efficiency percentage reported by Liu et al. (2011) and Ojaghian et al. (2018).
Southern blot test revealed that the lines TMB4, TMB7, TMB12, and TMB18 introgressed one, two, two, and two copies of the transgene, respectively (Fig. 3). Furthermore, no relation was observed between the level of chitinase activities and resistance to white mold. Takahashi et al. (2005) reported a positive relation between chitinase transcript accumulation and chitinase activity leading to higher resistance to disease. However, some studies (Emani et al. 2003; Nishizawa et al. 1999) showed no considerable relation between the chitinase activity and level of resistance against fungal phytopathogens.
Chitinolytic enzymes are classified into endochitinase and exochitinases such as ╬▓-N-acetylhexosaminidase and 1,4-╬▓-chitobiosidase (Duo-Chuan, 2006). Roberti et al. (2012) showed that the application of Trichoderma spp. increase both endochitinase and exochitinases in plan tissues. However, upregulation of chit42 in tuber mustard did not increase the activity of exochitinases in the transgenic plants. In the current study, growth parameters such as height, fresh weight, and dry weight of the treangenic plants were statistically similar (P < 0.05) to those of untransformed plants. After assessment of the growth factors in the lines TMB4, TMB5, TMB7, TMB12, TMB16, and TMB18, the mean values ┬▒ standard errors for height, fresh and dry weights of aboveground tissues in the transgenic plants were recorded as 25.2 ┬▒ 1.8 cm, 35.8 ┬▒ 3.2 g, and 8.9 ┬▒ 0.7 g, respectively. However, the mean values ┬▒ standard errors for the same growth factors in three untransformed controls were recorded as 24.8 ┬▒ 2.3 cm, 36.2 ┬▒ 1.2 g, and 9.3 ┬▒ 0.5 g, respectively. Further tests are carried out to assess the effect of introduced chit42 against white mold of tuber mustard under field condition in Zhejiang province. Introducing the gene chit42 to tuber mustard showed to be an effective method against white mold caused by S. sclerotiorum. It is suggested to test the effect of introducing chit42 to different cultivars of tuber mustard commonly grown in Zhejiang province. Further tests are going to be conducted to produce tuber mustard lines carrying a two gene construct including chit36 and chit42 from T. harzianum.



 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






