

Introduction
Eucalyptus was introduced into many countries worldwide including China due to its fast growth, high productivity and ever-increasing demand for paper, plywood, and fuel. Eucalyptus dieback disease is one of the most serious Eucalyptus diseases not only in China but also in many Eucalyptus plantation countries (Li et al., 2013). This disease is mainly caused by fungal pathogens of several species in Cylindrocladium (teleomorph: Calonectria) genus. For example, C. clavatum and C. quinqeseptatum were identified as the pathogens of Eucalyptus dieback in India (Sharma and Mohanan, 1982). Cylindrocladium and Cylindrocladiella spp. were found to be the pathogen of Eucalyptus dieback in Africa (Crous et al., 1993). According to a report by Crous and Wingfield (1994), C. quinqeseptatum caused Eucalyptus dieback in Australia, America and Brazil, and C. scopariuma also caused this disease in Spain, Thailand, Australia, and Brazil. Previous studies indicated that C. scoparium is one of the most important pathogens of Eucalyptus dieback and caused disease on Eucalyptus frequently in most parts of Eucalyptus planting areas in China (Zhu et al., 2011a). It was estimated that the Eucalyptus planting area reached more than 10 million hm2 in China, and 60% of which suffered Eucalyptus dieback infections in a certain degree. Eucalyptus dieback caused by C. scoparium has been defined as a key national plant quarantine disease in China (Pang et al., 2001). C. scoparium usually infects Eucalyptus seedlings and young Eucalyptus trees under 4 years old. In early stage of infection, this disease can easily lead to reddish-brown spots on leaves and shoots, leaves wilting, and branches dropping, which develop upwards from the base of young trees (Zhu et al., 2011b). In late stage of infection, it can cause large damage to the growth of Eucalyptus, result in branch wither, and lead to crown dieback or tree death depending on the severity of the disease (Li et al., 2013).
For a long time, the widest applied detection technique for Eucalyptus dieback disease is direct observation (Anonymus, 1984). However, this detection method by human judgment requires professional persons who have solid theoretical knowledge, and it is intensive time and labor consuming. Moreover, this method particularly has low levels of sensitivity and specificity, and it is hard to distinguish C. scoparium from other Cylindrocladium species. Therefore, it is not suitable for large scale diagnosis and surveillance. Although accurate detection of C. scoparium on Eucalyptus dieback can depend on pathogen isolation and classification by morphological, biological and genetic characteristics, it usually takes a long time to achieve a final result. To develop efficient strategies of fungal disease control as early as possible, a rapid and reliable detection technique is desired to accurately distinguish Eucalyptus dieback caused by C. scoparium from other Cylindrocladium species. However, such technique for detection of C. scoparium on Eucalyptus dieback has not been established till now. In recent years, some rapid molecular detection techniques, such as nested PCR, multiplex PCR and loop-mediated isothermal amplification (LAMP) have been successfully developed for detection of fungal diseases of plants (Grote et al., 2002; Guglielmo et al., 2007; Hansen et al., 2016).
The objective of this study was to develop a rapid, simple, easy operating, sensitive and specific method for detection of C. scoparium in most Eucalyptus materials. Here, we reported for the first time that three PCR-based methods, including nested PCR, multiplex PCR, and LAMP, were developed for detection of C. scoparium based on factor 1-alpha (tef1) and beta-tubulin gene. In particular, the sensitivity, specificity and field application of these new methods for detection of Eucalyptus dieback disease caused by C. scoparium were assessed.
Materials and Methods
Strains and culture conditions
The 22 fungal strains used in this study were listed in Table 1, including 10 Cylindrocladium isolates (6 isolates from C. scoparium and 4 isolates from other species in Cylindrocladium genus), and 12 none-Cylindrocladium isolates obtained from diseased leaves and stem tissues caused by Eucalyptus dieback, or healthy plants. These isolates were either available from Forest Protection of Chinese Academy (Beijing, China) or isolated and identified by sequencing and BLAST analyses of 26s and internal transcribed spacer (ITS), and deposited in the culture collection of Key Laboratory of Forest Protection of Sichuan Province, Sichuan, China. All isolates were stored on potato dextrose agar (PDA) medium covered by plum-extract agar with liquid paraffin at 4°C in the dark. Fresh cultures of isolates were prepared by placing small agar plugs from stock cultures in deionized water and then subculturing on PDA grown at 25°C. After 7-day incubation in dark, the mycelium was collected for DNA extraction or immediately stored at −20°C until use.
DNA extraction
Fungal mycelium and plant materials were ground and homogenized in liquid nitrogen using a mortar and pestle. The DNA was extracted from fungal mycelium and plant samples using a Plant Genomic DNA Extraction Kit (Tiangen Biotech Co., Ltd., Beijing, China) according to the manufacturer’s instructions. The DNA concentrations and purities were determined by a NanoDrop 2000C Spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA) and further verified by agarose gel electrophoresis.
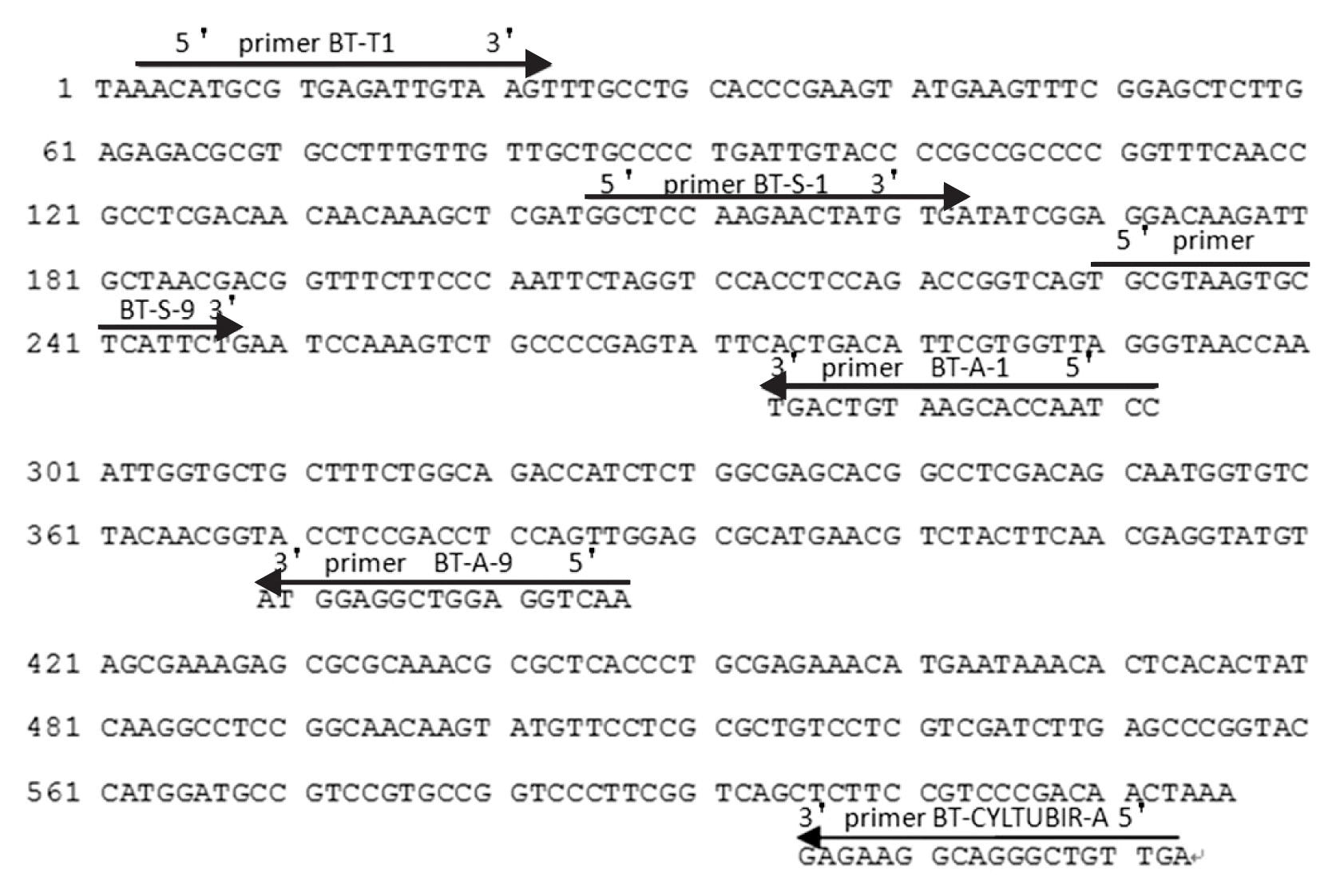
Fungal verification and primer design
All isolates used in this study were verified by amplification and sequencing of fungal ITS region using universal primer pair of ITS1 and ITS4 (Table 2) (White et al., 1990). Conventional PCR reaction was carried out in 50 μl of total volume consisting of 10 mM Tris-HCl (pH 8.0), 50 mM KCl2, 1.5 mM MgCl2, 0.3 mM each dNTP, 0.3 μM each ITS1 primer and ITS4 primer, 1.5 U Taq DNA polymerase (New England Biolabs, Beverly, MA, USA), and 4 μl of DNA template. The amplification protocol included an initial denaturation at 95°C for 5 min, followed by 30 cycles at 95°C for 30 s (denaturing), 55°C for 45 s (annealing) and 72°C for 90 s (elongation), and a final elongation at 72°C for 5 min. The PCR reaction was run on a Mastercycler Personal PCR Thermocycler (Eppendorf, Hamburg, Germany). All the PCR amplifications in this study were the same as the above unless described else. The PCR products were analyzed by 1% agarose gel electrophoresis. The beta-tubulin and factor 1-alpha (tef1) genes of Cyl-indrocladium genus were amplified and sequenced using universal primer pair BT-T1/BT-CYLTUBIR-A and EF1-728F/EF2 (Table 2), respectively (Hagemann and Rose, 1988). Based on the resulting sequences, nested PCR primers (BT-S-1/BT-A-1) (Table 2, Fig. 1) were designed in the beta-tubulin gene and multiplex primers (EF-S-4/EF-A-4 and BT-S-9/BT-A-9) (Table 2, Fig. 1, 2) were designed in the beta-tubulin and factor 1-alpha (tef1) genes using the Primer 6.0 (Premier Biosoft, Palo Alto, CA, USA). For LAMP assay, six primers, including two outer (F3 and B3), two inner (FIP and BIP) and two loop primers (LF and LB) (Table 2, Fig. 3), were designed using the Primer Explorer V4 (http://primerexplorer.jp/elamp4.0.0/index.html) as described previously (Mori and Notomi, 2009). All of the primers were synthesized by the Sangon Biotech Co., Ltd. (Shanghai, China).
PCR amplification and product detection
A nested PCR based on a two-step approach was developed in this study. In the first round of amplification, universal primers (BT-T1/BT-CYLTUBIR-A) were used to enrich the amount of fungal DNA present in all of the samples from the tested fungal isolates. The reaction system and amplification profile referred to the conventional PCR reaction as described in the previous section, except for the annealing temperature (58°C) and the primers (BT-T1/BTCYLTUBIR-A) (Glass and Donaldson, 1995; O’Donnell and Cigelnik, 1997). The second round PCR amplification was carried out by adding 10-fold diluted PCR product as template and BT-S-1/BT-A-1 as the primer pair.
For the multiplex PCR reaction, 1.0 μl of each primer (EF-S-4, EF-A-4, BT-S-9, and BT-A-9) was added into the amplification mixture and the annealing temperature was optimized by doing a temperature gradient from 51-59°C with 1°C increment.
LAMP reactions were carried out as described previously (Tomlinson et al., 2010). Briefly, a 50 μl reaction mixture contained 20 mM Tris-HCl (pH 8.8), 10 mM (NH4)2SO4, 10 mM KCl, 0.1% Triton X-100), 6 mM MgCl2, 0.2 mM each NTP, 0.3 M betaine hydrochloride (Sigma-Aldrich, St. Louis, MO, USA), 0.8 μM each internal primer (FIP and BIP), 0.1 μM each outer primer (F3 and B3), 0.4 μM each loop primer (LF and LB), 8 U Taq DNA polymerase (New England Biolabs, Beverly, MA, USA), and 4 μl of DNA template. The LAMP reaction mixture was incubated at 60°C for 60 min. Meanwhile, reaction without DNA template (supplemented with ddH2O to bring volume to 50 μl) was served as a negative control. Three different methods were used to examine the LAMP products, including visualization under ultraviolet (UV) light, on 2% agarose gel electrophoresis, and by fluorescent detection stained with SYBR Green I (Sigma-Aldrich).
Specificity and sensitivity of PCR primers
To evaluate the specificity of the designed primers, 22 DNA samples, including 6 DNA samples of C. scoparium, 4 samples from the same genus, and 12 samples from other 12 fungal genera (Table 1), were tested for nested-PCR, multiplex PCR, and LAMP assays. The sensitivities of nested-PCR, multiplex PCR, and LAMP were comparatively studied in this research. Briefly, the fungal DNA from pure culture of C. scoparium was used to make 10-fold serial dilutions (from 50 ng/μl to 500 ag/μl) to evaluate the sensitivity of these three assays in comparison to standard PCR assay. Regular PCR primed by primers of BTS-1/BT-A-1 were also carried out for comparative analysis. All of these three assays were conducted as described above on the serial dilution of DNA.
Detection of Eucalyptus samples
To evaluate the reliability of these three assays as a diagnosis system, we extracted the DNA samples from Eucalyptus plant in cultivation district, on June 2014. The collected samples included the fresh dead tissues, diseased tissues with typical symptoms, tissues infected at early stage, infected tissues, residues of dead plant, and healthy tissues. All these samples were tested by nested-PCR, multiplex PCR, and LAMP assay, respectively.
Results
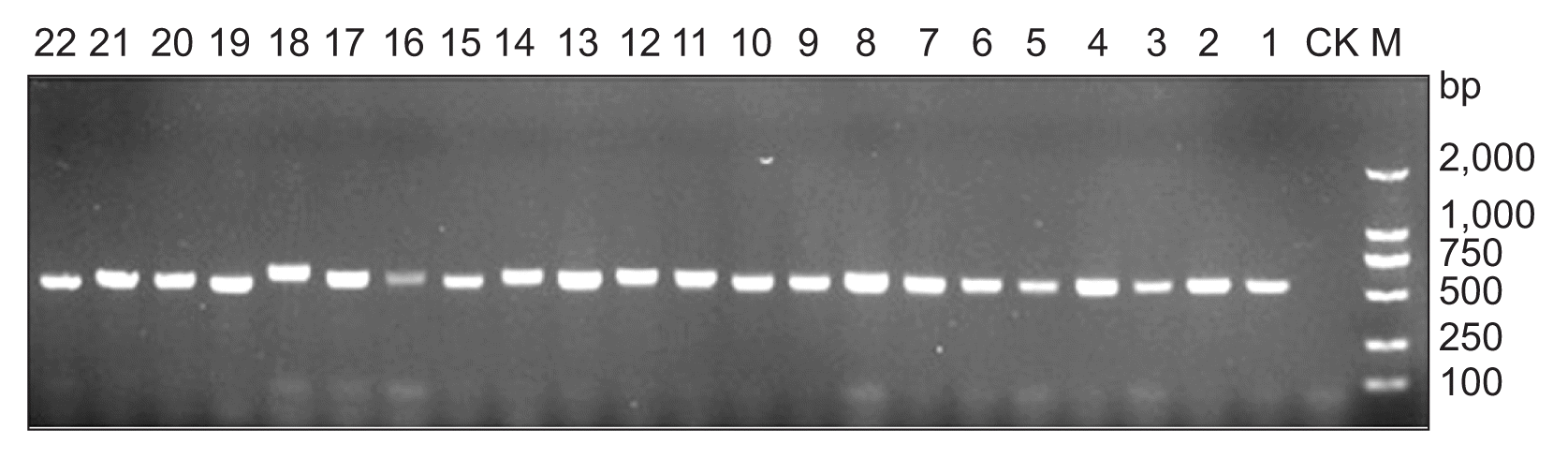
Evaluation of DNA quality. The concentrations of extracted fungal DNA from 22 fungal isolates were from 532 ng/μl to 675 ng/μl, and the ratios of A260/A280 of all the DNA samples were from 1.78 to 1.89. The quality of the extracted fungal DNA was further evaluated by amplification of ITS region using 50 ng of each extracted DNA as template and fungal universal primers of ITS1 and ITS4. Five Microliters of PCR products were used for agarose gel electrophoresis and the PCR products amplified from all the tested isolates displayed a single and bright band in each lane (Fig. 4), indicating the DNA quality was good enough for subsequent PCR reactions.
Specificity of designed primers
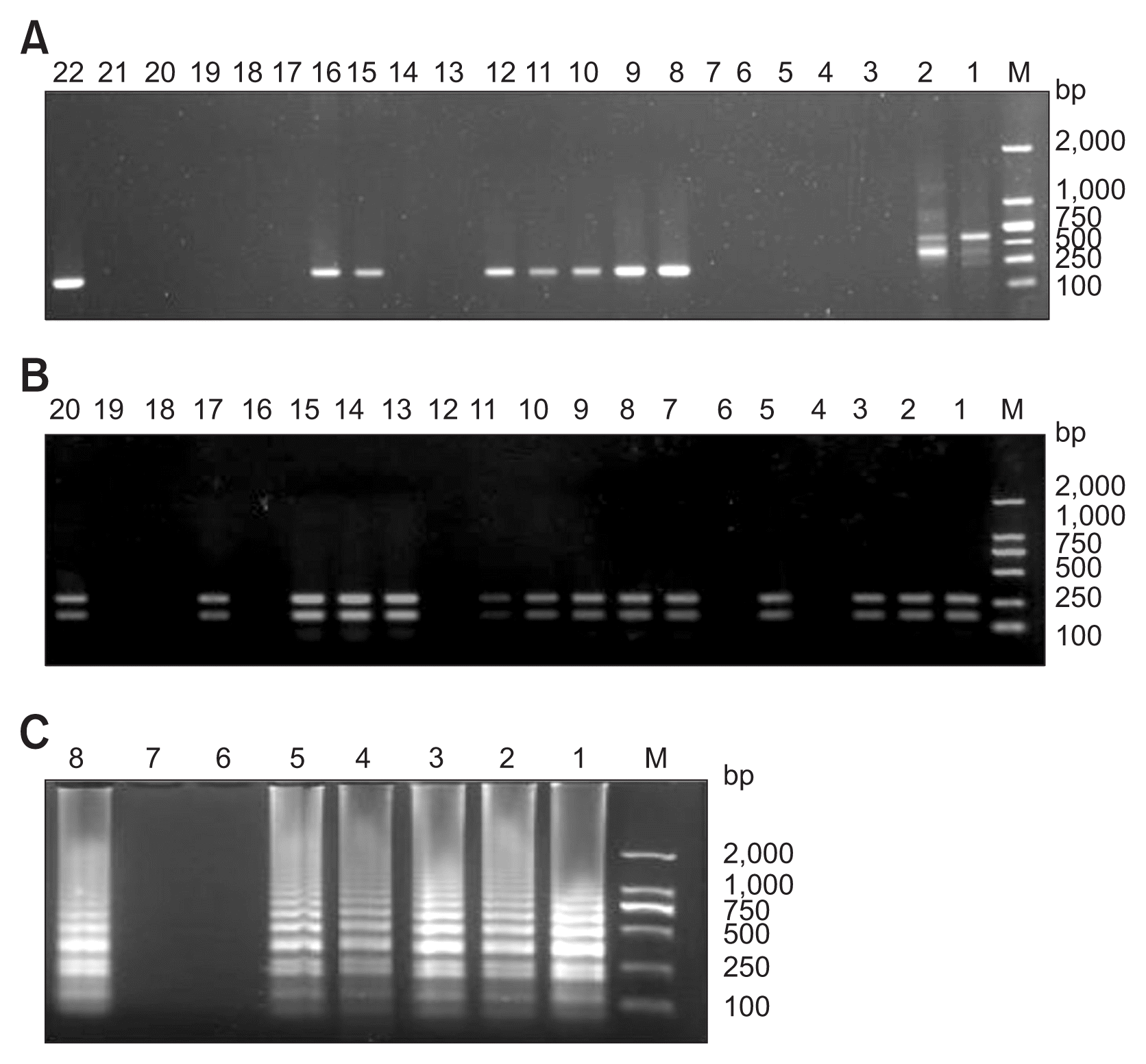
Using 50 ng of each extracted fungal DNA as template, an amplification product of the expected size of 148 bp was obtained with DNA from all the C. scoparium isolates by nested PCR (Fig. 5). Meanwhile, no amplification product was obtained from the negative control. Moreover, no amplification product was obtained with DNA from the isolates of other Cylindrocladium species and other genera tested as template in this study. Sequencing analysis indicated that the PCR product was the partial sequence of the beta-tubulin gene from C. scoparium. These results indicated that the designed primers for nested PCR are highly specific to C. scoparium.
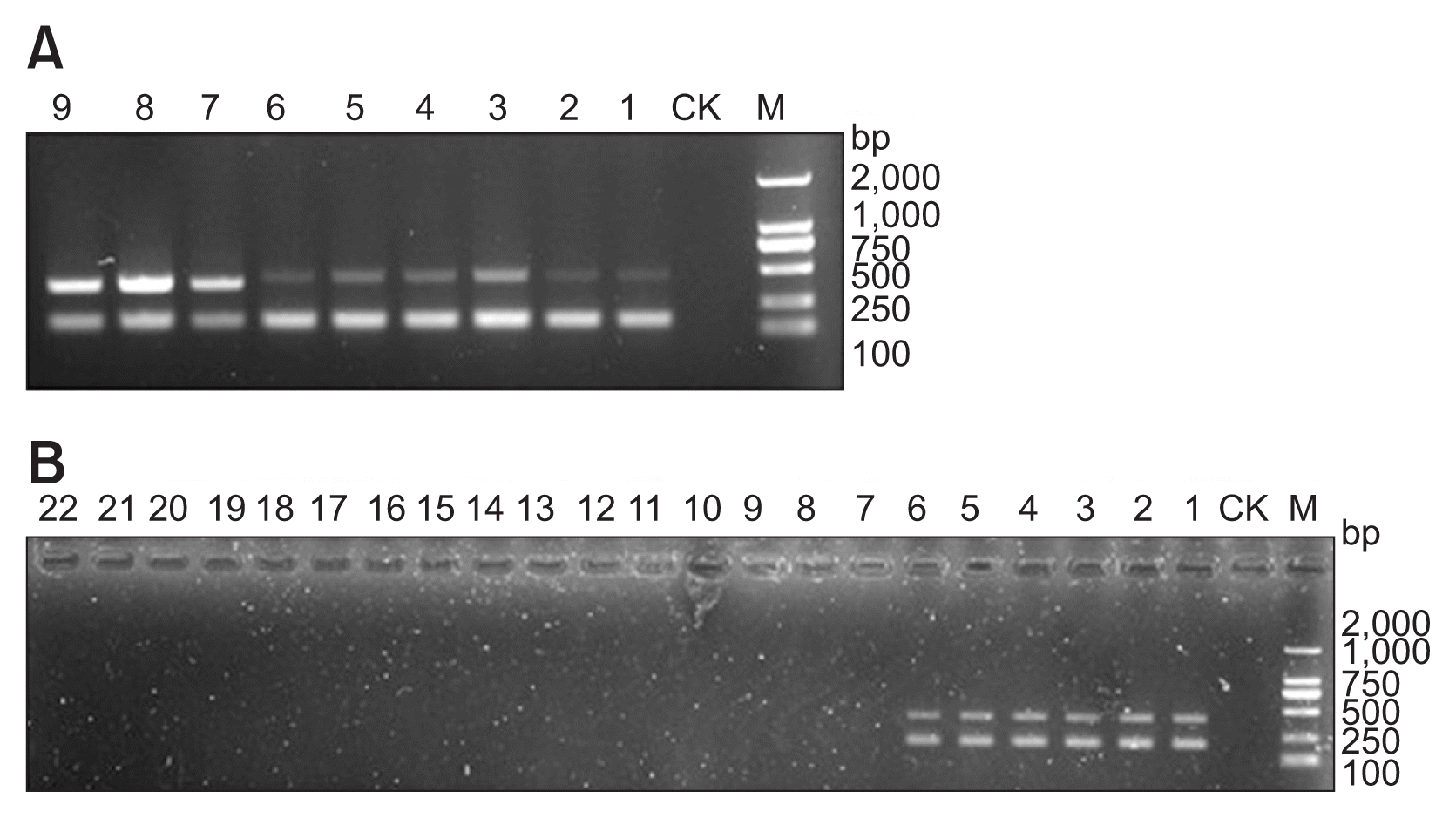
Optimization analysis of the annealing temperature of the multiplex method indicated that two clear bands with expected size (272 bp and 157 bp) were presented in the same lane on the electrophoresis gel at 58°C of annealing temperature (Fig. 6A). Good presentation of the two expected bands on the electrophoresis gel also indicated that there was no competition between the two primer pair for multiplex PCR amplification. Therefore, this temperature was used as annealing temperature of multiplex methods for later study and application. Specificity study of the designed primers for multiplex method showed that two amplification products were obtained with DNA from all the C. scoparium isolates as template (Fig. 6B). Like the nested PCR, no amplification product was obtained without DNA from the negative control, with DNA from the isolates of other Cylindrocladium species and other genera tested as template by multiplex PCR in this study. Sequencing analysis indicated that the two PCR products were the expected partial sequence of the beta-tubulin gene and tef1 gene from C. scoparium. These results indicated that the designed primers (EF-S-4/EF-A-4 and BT-S-9/BT-A-9) for multiplex PCR are highly specific to C. scoparium.
Optimal reaction for the LAMP assay with the primers sets designed in this study was found to be at 65°C for 45 min and ended at 90°C for 5 min. Using this optimized LAMP procedure, specificity of the primers for the LAMP was evaluated. Positive results were observed in LAMP assays with DNA from all the C. scoparium isolates as template under UV light, on agarose gel electrophoresis and by SYBR Green I fluorescent stain (Fig. 7). However, the LAMP reactions showed negative results without DNA from the negative control, with DNA from the isolates of other Cylindrocladium species and other genera tested as template by all of the three visualization methods in this study. Sequencing analysis indicated that the DNA products from LAMP assay were the expected partial sequence of the beta-tubulin gene from C. scoparium. All of these evidences indicated that the designed primers (F3/B3, FIP/BIP, and LF/LB) for LAMP assay are highly specific to C. scoparium.
Sensitivity of nested PCR, multiplex PCR, and LAMP
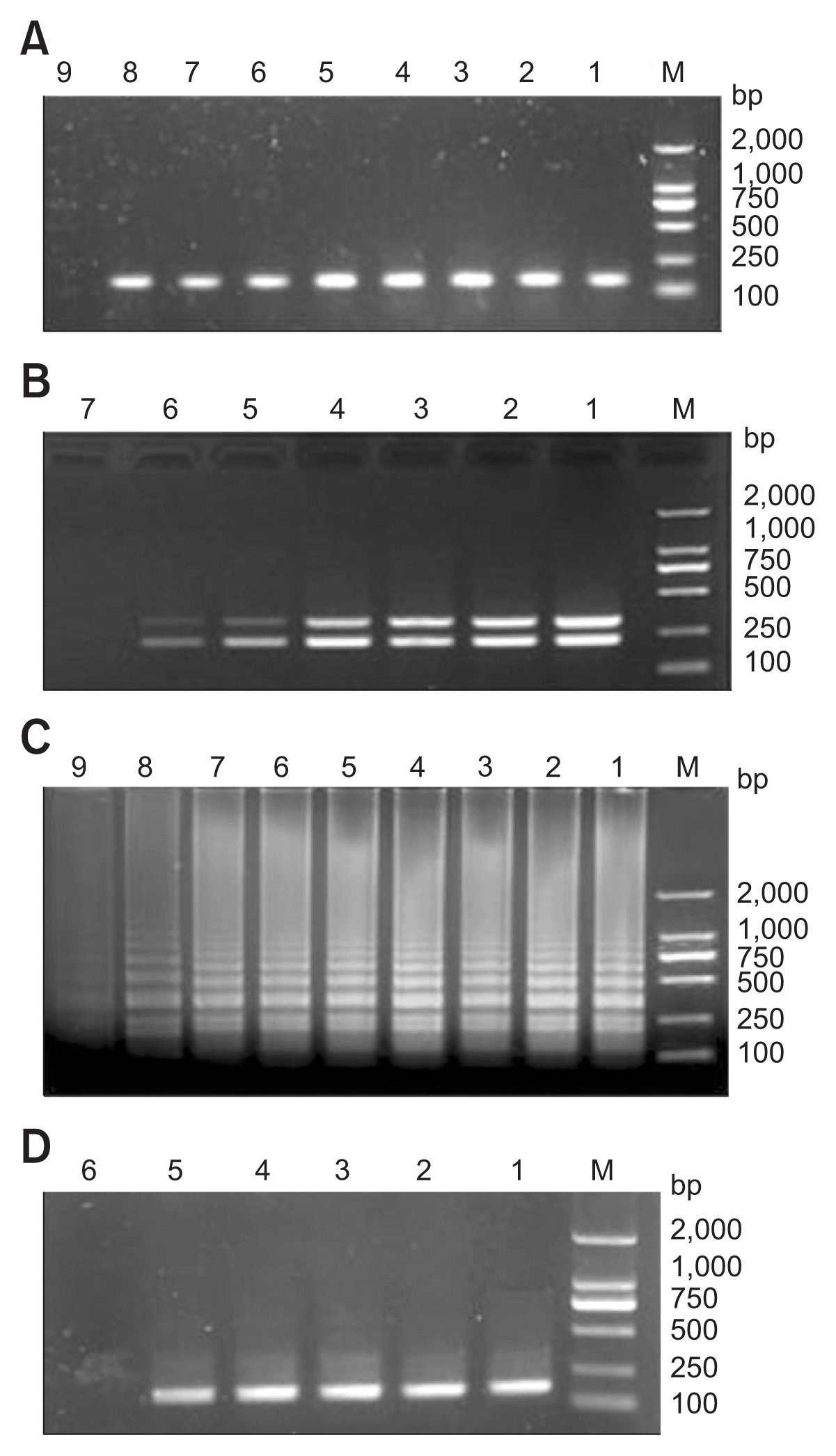
Sensitivity of nested PCR, multiplex PCR, and LAMP were detected on a serial 10-fold dilution of pure fungal DNA of C. scoparium (Table 3, Fig. 8). The nested PCR and LAMP showed much higher sensitivity than the multiplex PCR developed in this study. The detection limit of these two PCR-based methods was approximately 5 fg/μl of pure fungal DNA (Fig. 8A, C), which was much higher than the regular PCR (5 pg/μl) (Fig. 8D) by 1,000 folds. Although the sensitivity of the multiplex PCR (50 fg/μl) (Fig. 8B) was lower than that of the nested PCR and LAMP, it was 10-fold higher than the regular PCR (Fig. 8D).
Detection of Eucalyptus samples
Using regular PCR with primers of BT-S-1/BT-A-1, C. scoparium was detected in fresh dead Eucalyptus tissues and diseased tissues with typical symptoms (Fig. 9A). In comparison, C. scoparium was detected not only in the two tissues mentioned above but also in tissues infected at early stage, infected tissues, and residues of dead plant by nested PCR and LAMP (Fig. 9A, C). Similar to nested PCR, C. scoparium was also detected in most tissues mentioned above except for some samples from infected tissues by multiplex PCR (Fig. 9B). Considering LAMP assay’s characteristics of one-step and rapid reaction, simple operation, single-tube utilization, and simple visualization of amplification products, it was the most practical method for detection of C. scoparium.
Discussion
Eucalyptus dieback can be caused by several rather than one species in Cylindrocladium genus (Crous et al., 1993). It is very important to accurately detect the pathogen of dieback for later efficient control plan-making. Therefore, specificity of the detection method should be considered first in disease diagnosis (Rees-George et al., 2010). To well distinguish C. scoparium from other species in the same Cylindrocladium genus, representative isolates from four other species (C. colhounii, C. gracile, C. parasiticum, and C. quinqueseptatum) in Cylindrocladium genus were used as test strains in this study to evaluate whether the new developed nested PCR, multiplex PCR and LAMP are specific to C. scoparium. In addition, strains from 12 fungal genera usually isolated from Eucalyptus were included in this study. Some of them are pathogens to Eucalyptus, causing anthrax, peliosis, canker, grey speck, or round spot symptom, which undoubtedly will influence diagnostic results by direct observation. Therefore, using as many as possible strains of different species in the same genus and different genera from Eucalyptus as controls in the study could make the diagnosis of C. scoparium more accurate. To eliminate the influence of geographic variations on diagnosis by nested PCR, multiplex PCR and LAMP, we used six C. scoparium strains isolated from Sichuan, Guangxi, Fujian, Guizhou, Chongqing, and Guangdong provinces in China as representative strains.
A strongly conservative and species-specific sequence is the key factor to design primers successfully (Bonants et al., 1997). Beta-tubulin gene (also called β-tubulin gene), was widely used in the classification, identification, and phylogenetic analysis of fungi, bacteria and actinomycetes etc., due to its well specificity (Tan et al., 2009). Similarly, factor 1-alpha gene, existed both in prokaryotes and eukaryotes, was also proved to be specific to species (Huang et al., 2007). In this paper, we designed the nested PCR and LAMP primers based on the sequence of beta-tubulin gene, and multiplex PCR primer based on the sequences of both beta-tubulin gene and factor 1-alpha gene. Specificity studies established that all the primers used for nested PCR, multiplex PCR and LAMP were well species-specific to C. scoparium. In general, multiplex PCR is used to diagnose various species of pathogens. However, we developed this method for detection of C. scoparium in order to improve the diagnostic specificity in this study. Based on this foundation, a more complicated multiplex PCR for diagnosis of main species causing Eucalyptus dieback should be developed in further study.
The optimized reaction system was an essential prerequisite for the quantity of amplification products (Li et al., 2008). Previous study proved that an appropriate annealing temperature was as important as an optimized reaction system (Sipos et al., 2007). The annealing temperature of these three detection methods was optimized, respectively, in this study. The best temperature for nested PCR and multiplex PCR was at 58°C, while for LAMP it was 65°C.
PCR-based techniques, such as regular PCR, nested PCR, multiplex PCR, and LAMP, have been widely used for diagnosis and monitoring of plant diseases caused by microorganisms (Feng et al., 2012; Grote et al., 2002; Henson and French, 1993; Wang et al., 2010). Compared with traditional or conventional method (such as direct observation, pathogen isolation and identification, etc.) for detection of C. scoparium, the PCR-based methods possess some advantage characteristics of rapid, simple, reliable detection. However, there was no report of PCR-based diagnostic techniques covering the nested PCR, multiplex PCR, and LAMP on detection of C. scoparium. In this study, we developed these three methods for detection of one kind of pathogen at the same time, and compared them to regular PCR and within themselves using the same research materials and laboratory conditions. The comparative results based on data from such conditions are more reliable than other reported results based on data from different research groups and geographic regions. In this study, we found that the sensitivities of the nested PCR and LAMP were similar and higher than multiplex PCR, and the latter was also higher than regular PCR. Using different Eucalyptus tissues as samples for C. scoparium detection, all of the three PCR-based methods showed much better detection results than regular PCR. Considering the good sensitivity, the nested PCR and LAMP developed in this study are the priority selection for the specific detection of in laboratory diagnosis. Moreover, LAMP is the most practical method in field application due to its one-step and rapid reaction, simple operation, single-tube utilization, and simple visualization of amplification products. In order to achieve this goal of C. scoparium detection in field, a Rapid Fungi Genomic DNA Isolation Kit (Sangon Biotech Co., Ltd.), which lyse cells by heat, can be used for fungal DNA extraction, and a portable electric generator and a metal-bath heater need to be equipped.
In conclusion, any of the new developed nested PCR, multiplex PCR, and LAMP is a rapid, sensitive, and reliable diagnostic method for detection of C. scoparium on Eucalyptus dieback disease. All of these three PCR-based methods can be used as valuable tools in the indoor diagnosis of Eucalyptus dieback disease caused by C. scoparium, and LAMP can also be used as valuable tool in outdoor diagnosis. This study can contribute to the efficient prevention and control of Eucalyptus dieback disease caused by C. scoparium.




 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






