

Introduction
Tan spot caused by the fungus Ascomycetes/Dothideomycetes Pyrenophora tritici-repentis is a serious disease of wheat. The fungus has a wide host range including numerous non-cereal grasses (Krupinsky, 1992; Morrall and Howard, 1975). It is one of the pathogen associated with leaf spot diseases of wheat. The disease is on increase in both incidence and severity on wheat grown in the Mediterranean region (Benslimane et al., 2006, 2011; Nsarellah and Mergoum, 1997). The initial infection occurs through the ascospores that infect and produce lesions on young wheat seedlings (Adee and Pfender, 1989; Howard and Morrall, 1975; Wiese, 1987). The initial symptoms of tan spot are small dark brown spots that expand to become tan diamond-shaped lesions with a yellow halo (Marshall, 2009). Frequently, there is a small black spot in the center of the lesion. The lesions often coalesce as they grow, resulting in large necrotic area and hence reduces the photosynthetic area.
The etiology, biology and epidemiology of the fungus have been studied extensively by several researchers (Adee and Pfender, 1989; da Luz and Bergstrom, 1986; Hosford, 1971; Howard and Morrall, 1975; Morrall and Howard, 1975; Rees et al., 1982; Sutton and Vyn, 1990; Wright and Sutton, 1990). Variability among P. tritici-repentis populations was demonstrated by several workers around the world (Ali and Francl, 1998; Ali et al., 2010; Benslimane et al., 2013; da Luz and Hosford, 1980; dos Santos et al., 2002; Friesen et al., 2005; Lamari and Bernier, 1989, 1991; Leisová et al., 2008; Misra and Singh, 1972; Moreno et al., 2008; Schilder and Bergstrom, 1990; Singh and Hughes, 2006). Studying variability within the population in a geographical region is important because it documents the changes occurring in the population. A number of authors have studied monospore isolates of P. tritici-repentis their biology, morphology and culture peculiarities on different nutrient media and found variation among the isolates (Ali and Francl, 1998; Friesen et al., 2003; Hunger and Brown, 1987; Mielke, 1999; Wolf, 1991; Wolf and Hoffmann, 1993). A little or no information is available on P. tritici-repentis populations present on wheat in various geopgraphical regions of Algeria. This would be a prerequisite for a further population analysis of pathogen virulence and that ultimately would help in evaluation wheat cultivars for tan spot resistance grown in Algeria.
The objective of the present study was to characterize and compare several monospore isolates of P. tritici-repentis, collected from different infected wheat fields from various locations of Algeria, to find the morphological culture differences among them, if any.
Materials and Methods
Collection of isolates
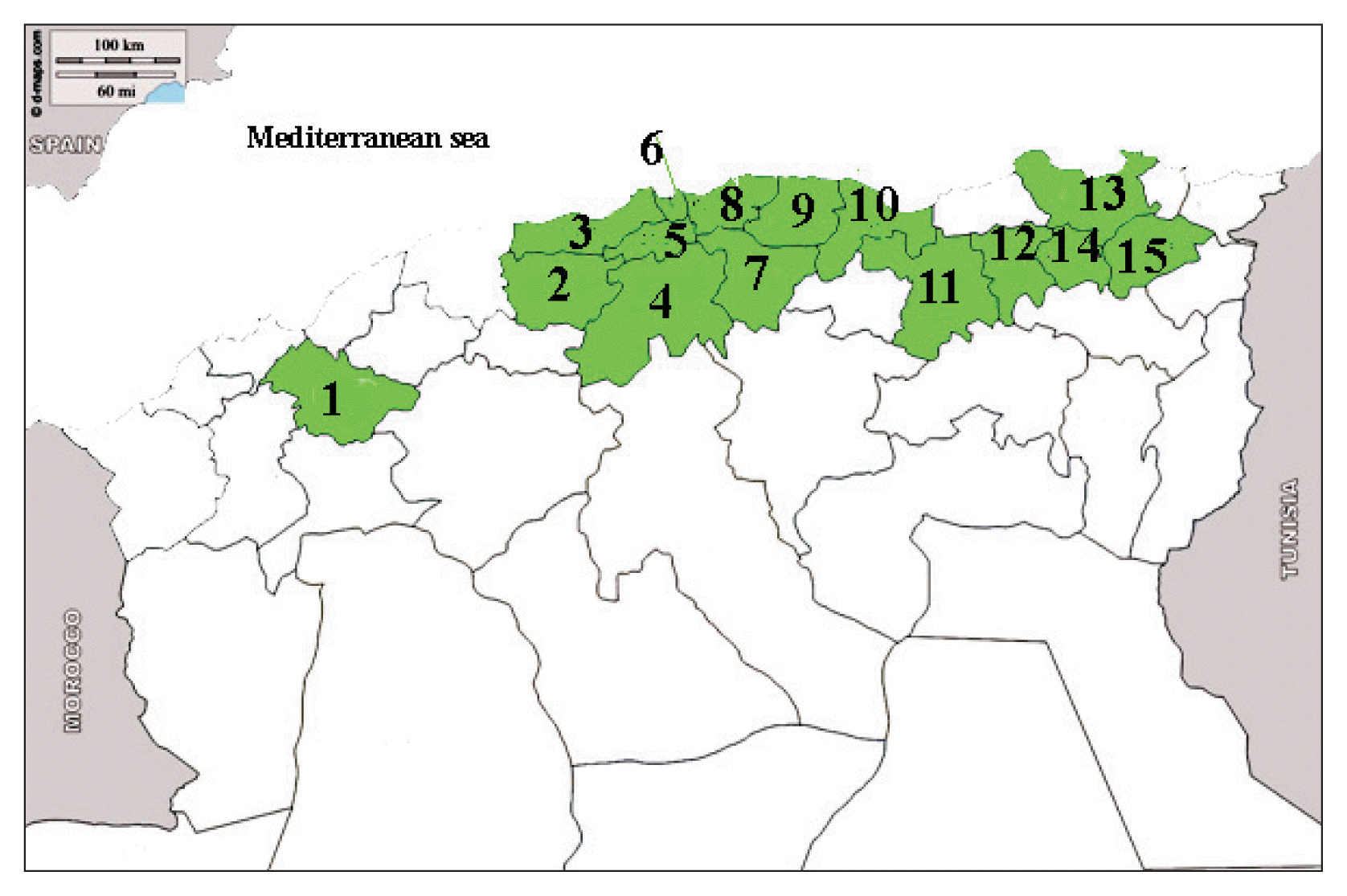
In a previous study (Benslimane et al., 2011), eighty-two mono-conidial isolates of P. triticirepentis were recovered from Triticum aestivum and T. durum diseased leaves sample. The samples were collected from different geographical wheat growing regions of in Algeria (Table 1, Fig. 1). Briefly, leaf spotted area were cut into 3 cm pieces, surface sterilized in 5% hypochlorite solution for 3 min, then rinsed thrice in sterile water (5 min each time). The fragments were blotted on tissue paper to remove the excess water and placed in Petri dishes with three layers of dampened Whatman filter paper. The plates were incubated at 22°C for 24 h under light then in dark for 24 h. Single conidia from conidiophores (Fig. 2) developing close to the edge of each lesion were transferred to potato dextrose agar (PDA).
Colony texture and color determination
All 82 isolates were grown individually on PDA in plastic Petri dishes at 20°C in the dark. After one week, 5 mm diameter plugs were taken aseptically from the margins of actively growing cultures and placed fungus-side down in the center of fresh PDA dishes. Each isolate was replicated 4 times. Petri dishes were incubated in dark at 20°C, and then macroscopic characters (color, sector, and texture) of each colony were recorded after 7 days. PDA was choosing for this step and the next, because it is commonly used for the isolation and growth of wide range of fungi in laboratories.
Mycelial radial growth evaluation
All isolate hyphal growths were determined on PDA medium. To assess the effects of temperature on in vitro colony radial growth, small plugs, 5 mm in diameter were transferred singly to 9 cm Petri dishes containing PDA. Cultures were incubated in the dark at three temperatures (20°C, 25°C, and 30°C), mycelium radial growth was recorded for each isolate in mm at 24 h interval until the colonies had reached the plate edge. Four diameters of linear growth of each plate were measured at right angles to each other and the values were averaged. Four replications (one plate/rep) were used for each isolate and for each temperature. Means values were used to perform principal component analysis using STATISTICA software (StatSoft Inc., Tulsa, OK, USA).
Conidia characteristics identification
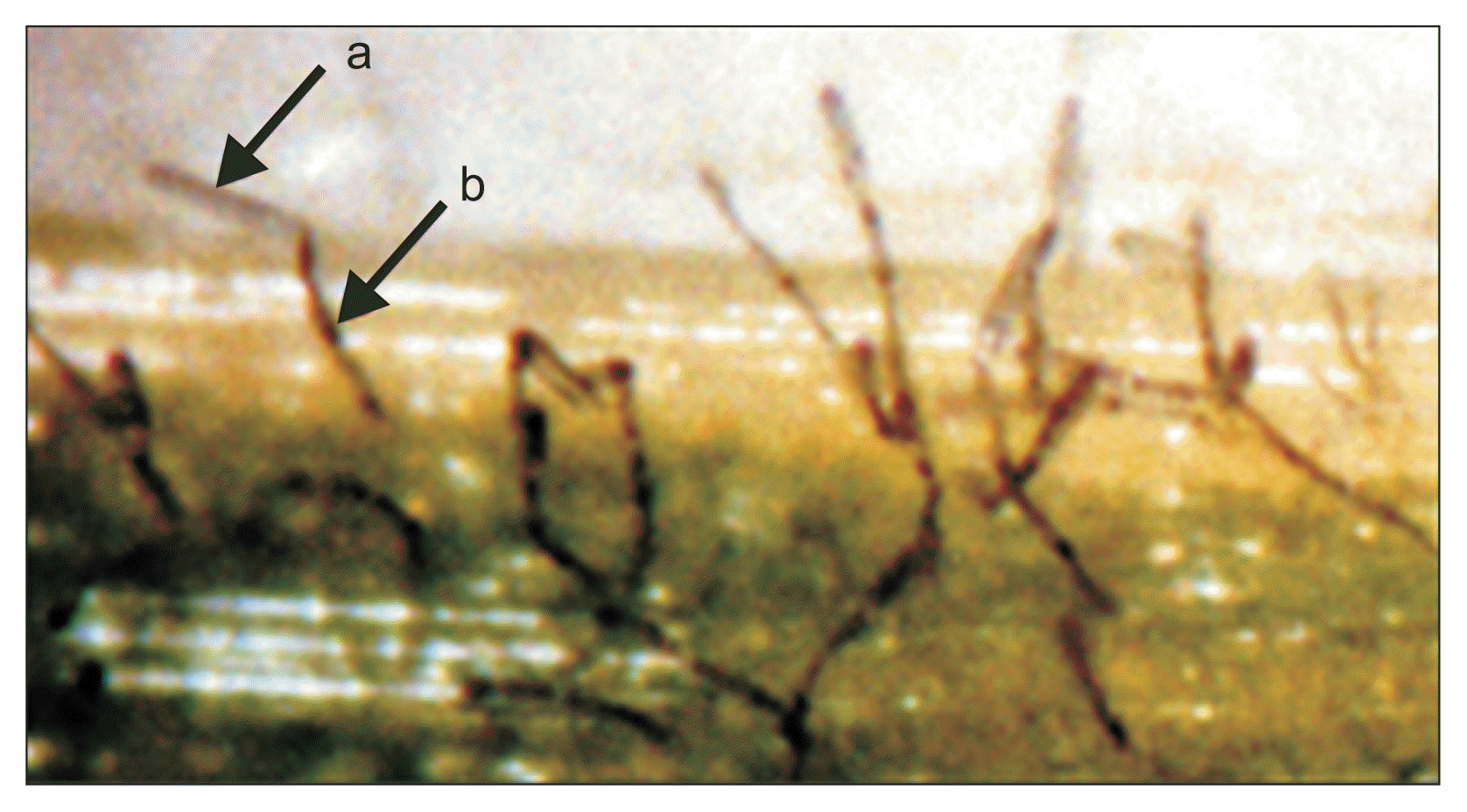
To determine whether any differences in conidial width and length exist within or among the isolates, 42 isolates were selected randomly and spores were produced of each isolate following the protocols of Lamari and Bernier (1989). Small plugs, 0.5 cm in diameter, from 8 day culture were transferred singly to 9 cm Petri plates, containing V8-PDA. The cultures were incubated in the dark until the colonies reached 4 cm in diameter. The cultures were then flooded with sterile distilled water and the mycelium flattened with the bottom of flamed test tube. After the water was decanted, the cultures were subjected to a regime of 18 h of light at room temperature followed by 24 h of dark at 15°C. Conidia size (length, width) and septation number of 50 spores/each isolate were counted under 40× magnifications with the aid of ocular and stage micrometer in compound microscope. Spore measurements were compared among the isolates and data were analyzed statistically by ANOVA method using SPSS software (SPSS Inc., Chicago, IL, USA).
Results
Colony texture and color
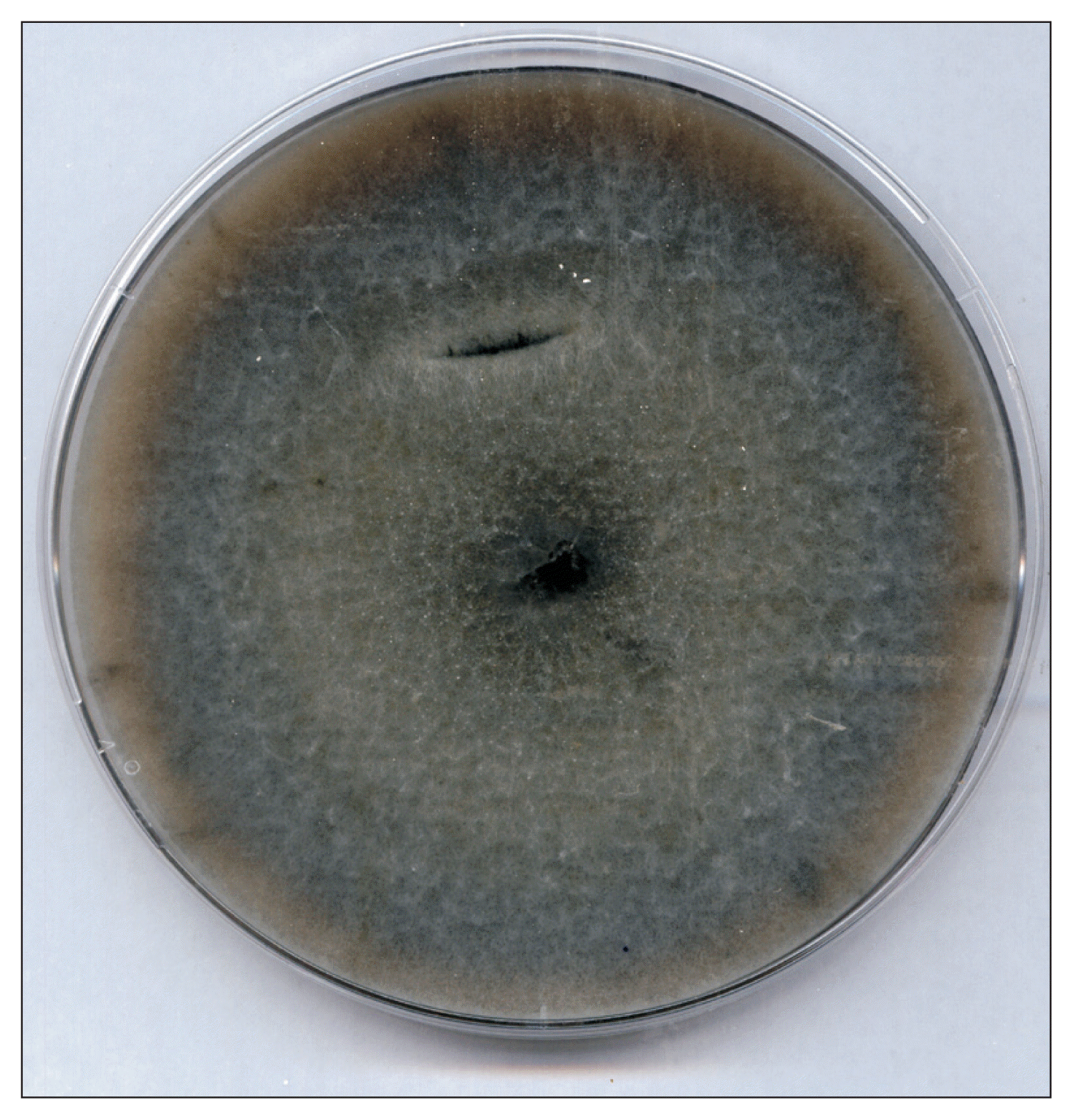
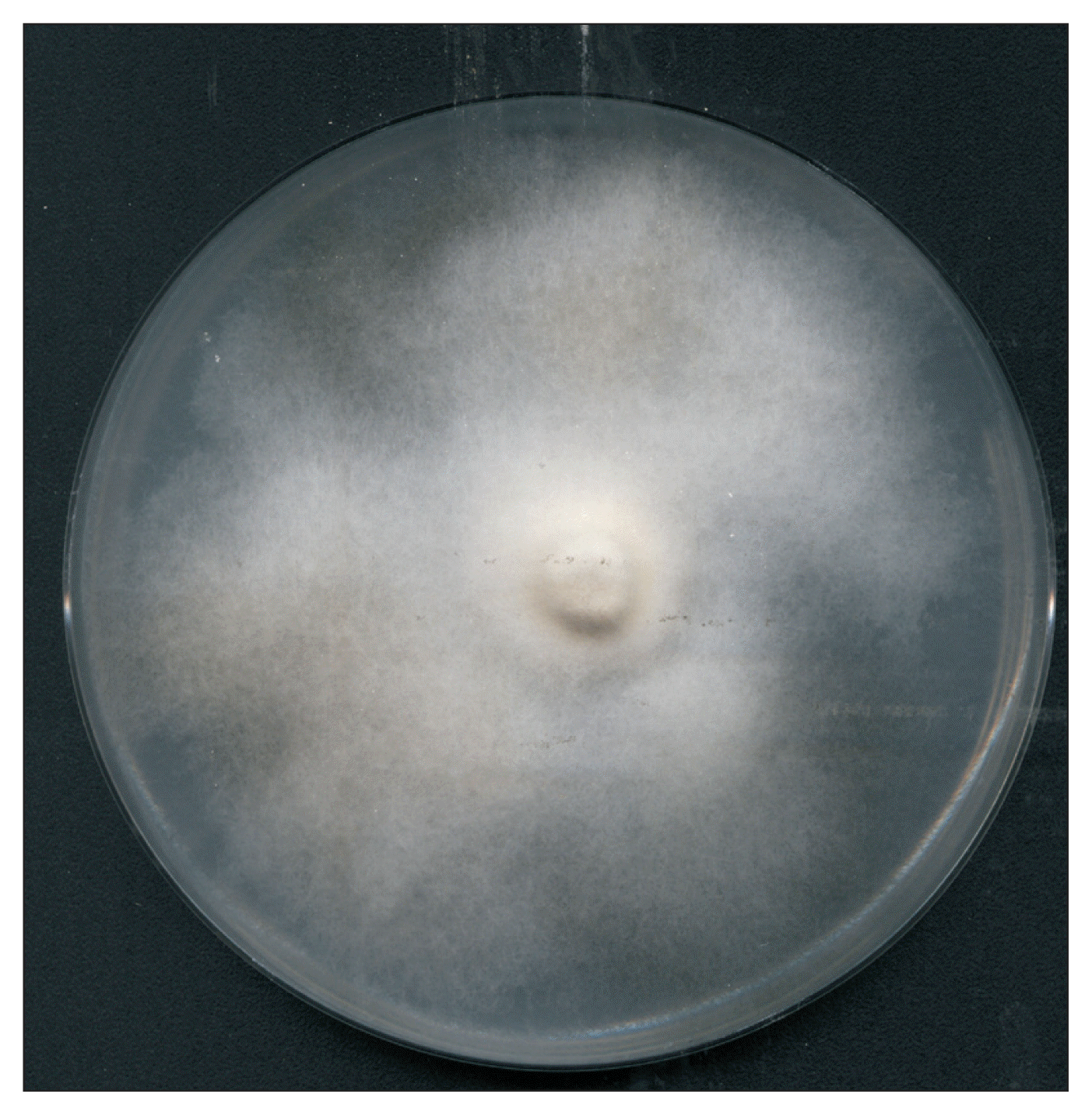
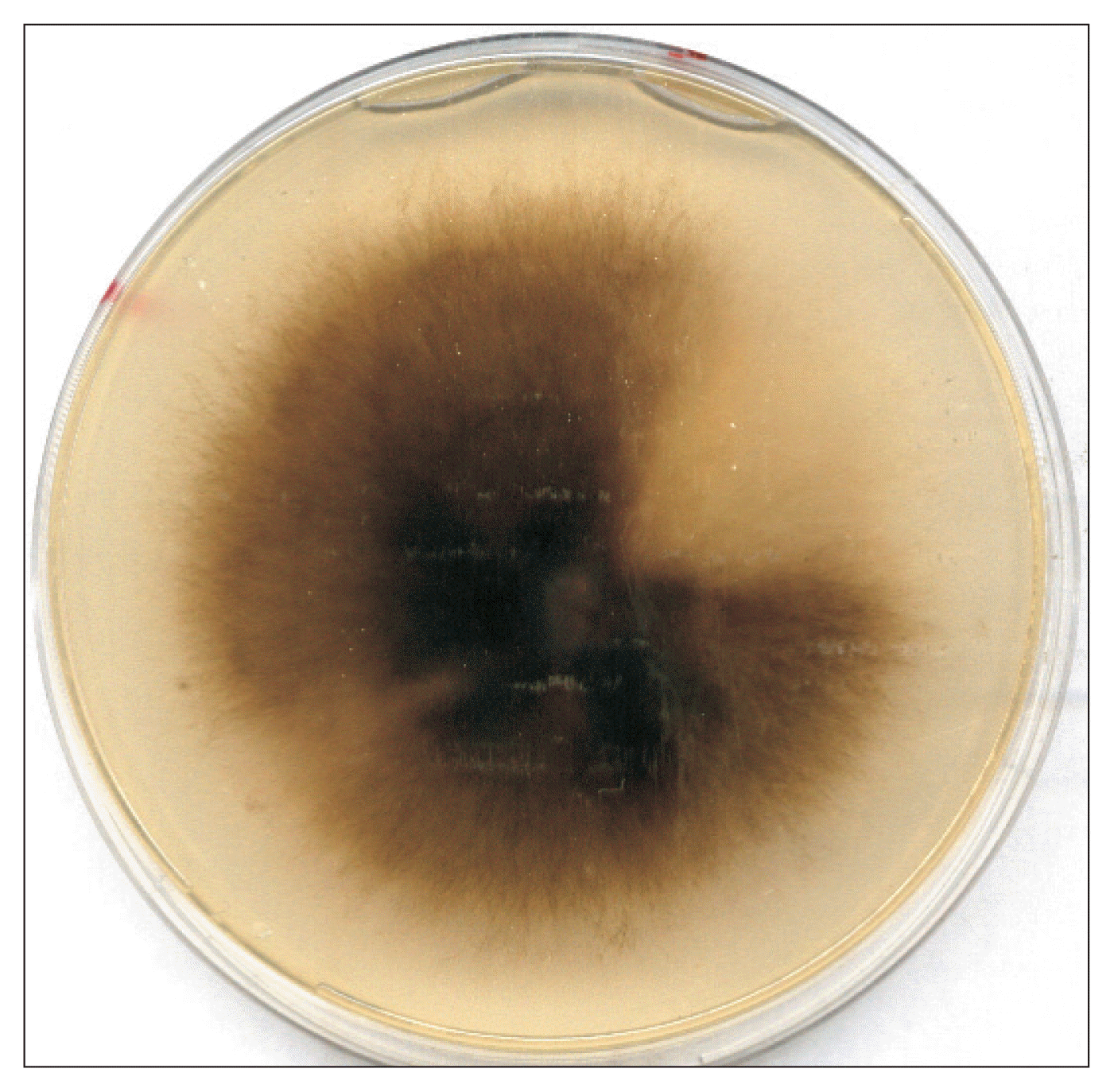
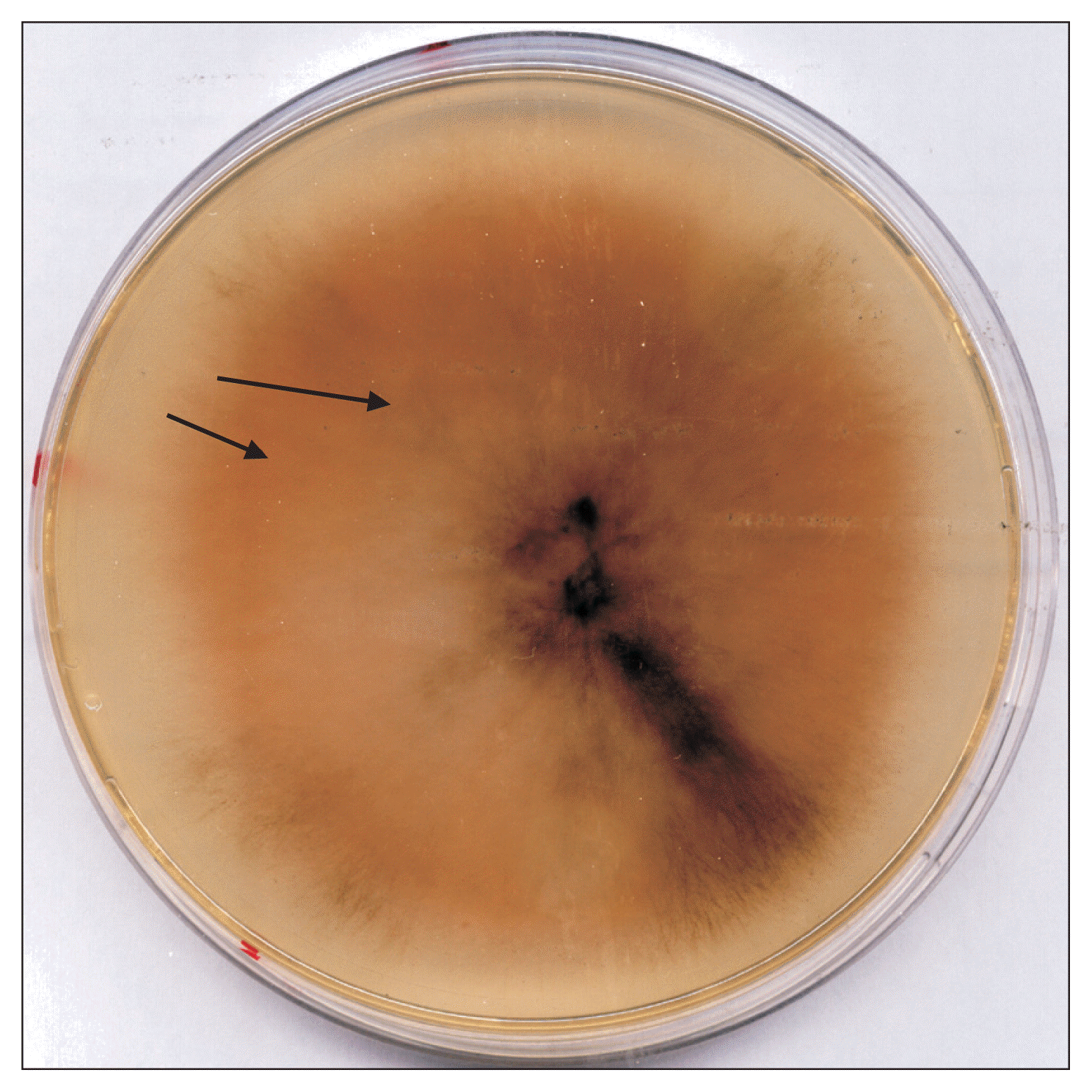
The P. tritici-repentis isolates used in this study varied in colony color and mycelium compactness. Cultures growth on PDA medium showed usually a thick cottony mycelium sometimes fluffy, often gray-green (Fig. 3), and rarely whitish (Fig. 4). Underside the colony color was green (Fig. 5). The old cultures about two weeks showed black spherical mycelia aggregations (Fig. 6). Moreover we found that when the mycelium color became orange, the isolates loosed the ability to produce spores as a result of a repeated sub-culture in PDA medium (Fig. 7).
Mycelial radial growth
The results on radial growth of all isolates colonies at different temperatures showed that the mycelium growth is temperature depended (Table 2). It was observed that the temperature range of 25-30°C was optimum for mycelial growth of P. tritici-repentis on PDA. Indeed, most of the isolates (57) showed a better (3.72-6.95 mm) growth at 25°C; however, some (10) of the isolates grew better at 30°C. Only five isolates (Ptr16, Ptr20, Ptr41, Ptr48, and Ptr72) grew (3.45-5.55 mm) better at 20°C. Based on radial growth at the three different temperatures, the principal component analysis categorized the isolates into six groups (Fig. 8); some of these groups stand out more than others; such as the group consisting of a single isolate Ptr6, or the group combining Ptr52 and Ptr64 (Table 1).
Even the different analyses of the results of mycelia growth showed that there was some difference between the studied isolates, the effect of temperatures, expressed trough the radial growth, showed that the difference or the approximation among studied isolates, seems to have no relationship with the geographical origin of the isolates. It was found that pathogen population in closely located fields or in the same, consisted of specimens of large phenotypical variability. Otherwise, it does not show any relationship with the climate of the area from where the sample was collected. In deed in Fig. 8, we can see isolates from different geographical regions grouped together, as well as other isolates from nearby areas classified statistically in groups far away each other.
Conidia characteristics
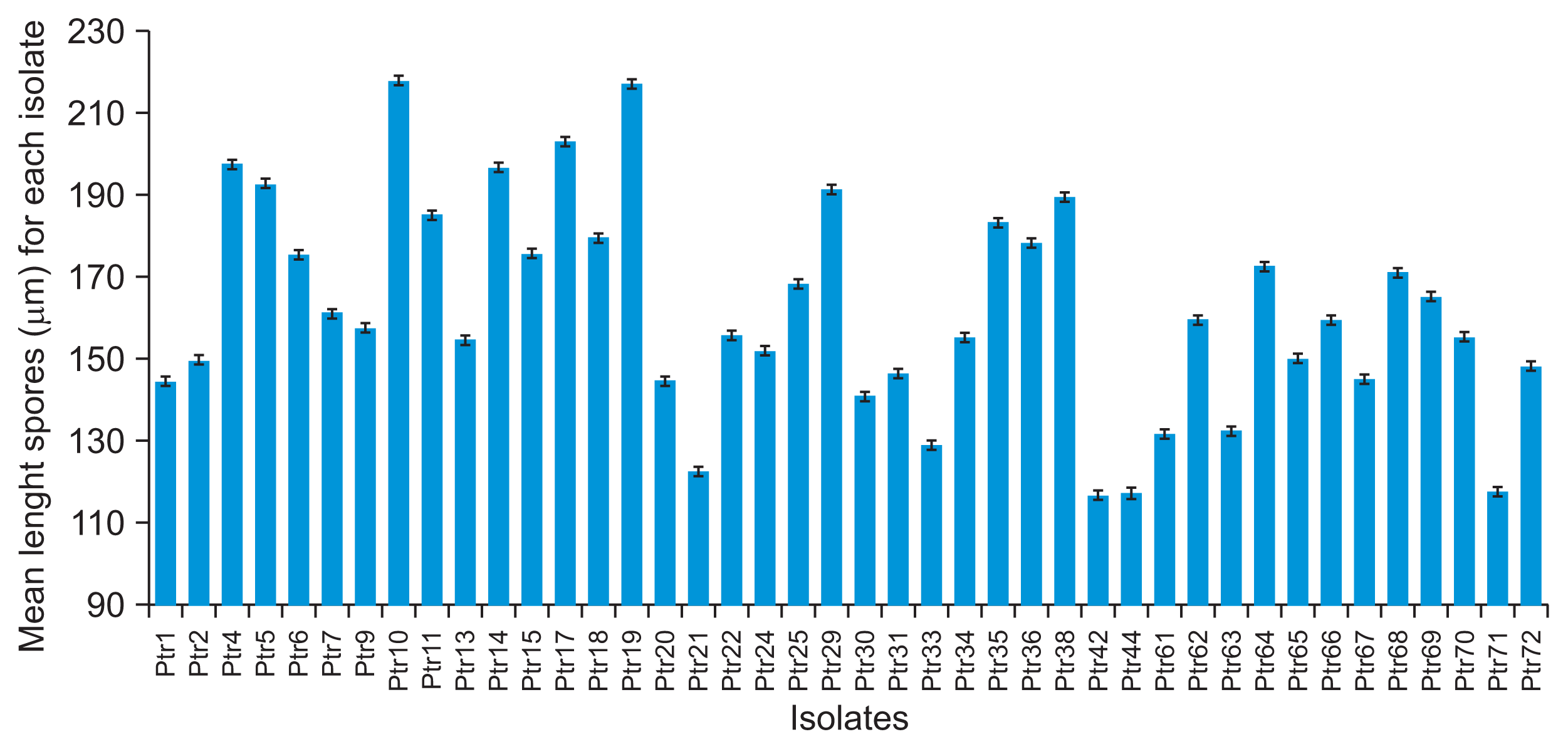
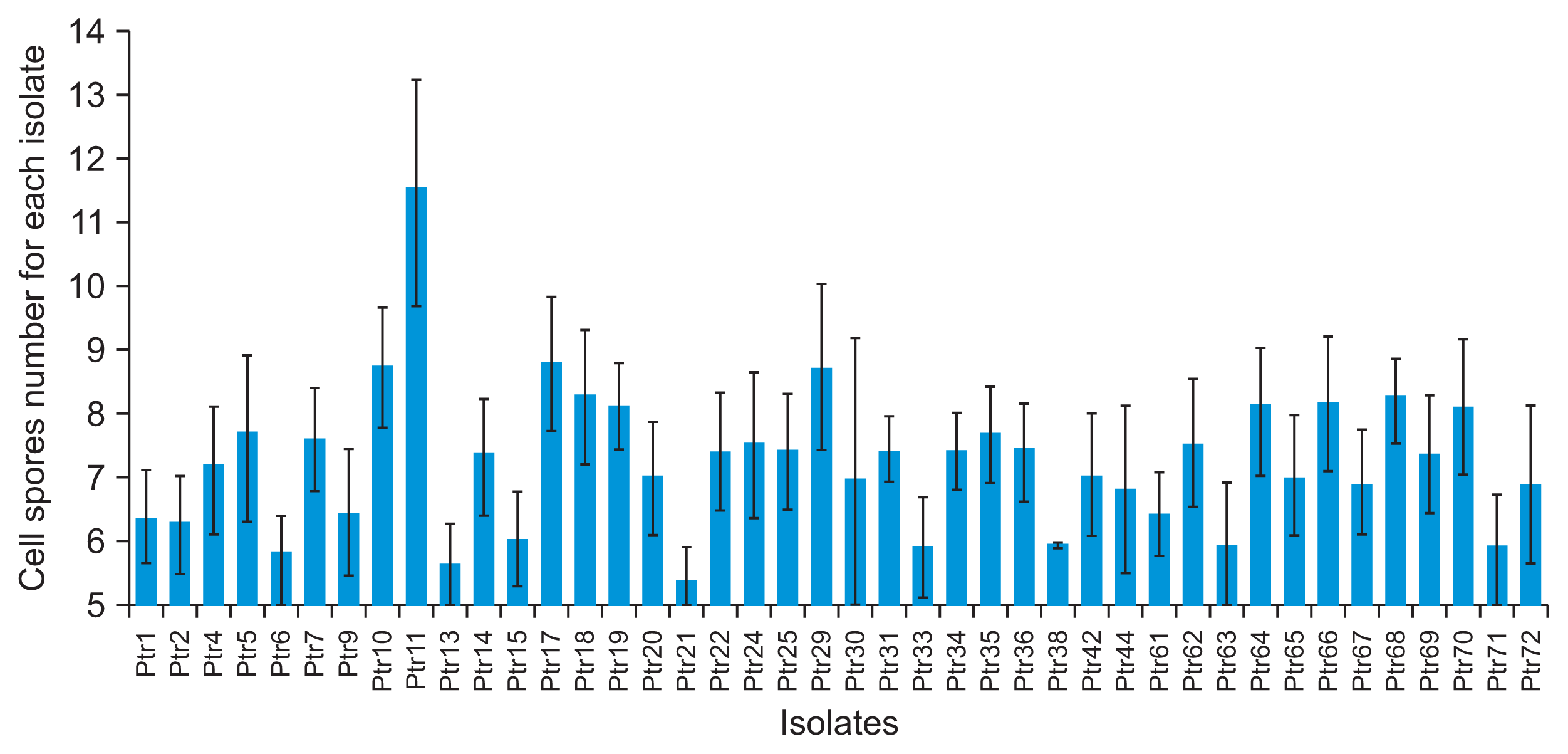
P. tritici-repentis isolates produced abundant conidia on V8-PDA which were consistent in morphology. Significant differences were founded in conidial length and number of cells among the isolates (P = 0.000). Dimensions of the conidia for each isolate are presented in Fig. 9 and Fig. 10. Average conidial length was maximum (217.67 μm) in isolate Ptr10 and minimum (117.15 μm) in Ptr71 whereas the number of septa varied from 11.52 in isolates Ptr11 to 5.42 in isolate Ptr21. However, no significant variation was observed in conidial width; only two values were founded 15.6 μm and 18.2 μm.
Discussion
The results of the present study revealed wide morphological variation among P. tritici-repentis isolates based on colony color and texture, mycelial radial growth, and conidial size prevalent on wheat in Algeria.
Several researchers observed variation in mycelia color and colony morphology in P. tritici-repentis and its related species isolates collected from different geographical regions. Dos Santos et al. (2002) studied P. tritici-repentis, McDonald (1967) and Frazzon et al. (2002) studied Pyrenophora teres isolates, and observed significant morophological variation based on mycelial colony colors. Similar results were reported by Benslimane (2002) and Christensen and Graham (1934) when they studied Pyrenophora graminea isolates for their morphological variation. Hosford (1971) observed that P. tritici-repentis isolates lose their sporulation when the culture became orange colored due to frequent subcultuing on PDA. This phenomenon also occurred frequently as sectoring in an otherwise typical colony and was not always associated with slower growth.
In Bipolaris sorokiniana isolates, Valim-Labres et al. (1997), Oliveira et al. (1998), and Matsumura (1991) observed and reported variability based on mycelium color and colony morphology grown on PDA. Some isolates exhibited white tufts while others showed fan shaped sectors, although the surface of all isolates was plain.
Conidia of Drechslera tactylidis significantly vary in length, diameter and number of septations (Zeiders, 1980). This also holds true for the closely related specie B. sorokiniana, when 87 isolates representing different agroecological regions of Pakistan studied for morphological variation and observed differences in colony size and conidial color and size (Asad et al., 2009).
Morphological variability is also common in several other plant pathogens population. In Fusarium oxysporum f.sp lentis, 32 isolates collected in western Algeria showed variability in the cultural colony appearance and size of conidia (Belabid, 2002). Similarly, 29 isolates of Sphaeropsis sapinea from Canada revealed several morphotypes based on their appearance of colony, their radial growth, and conidial size (Hausner et al., 1999).
Morphological variation within a taxon is well known in fungi (Harrington and Rizzo, 1999). For example it is known that the morphology of conidiophores and conidia in several asexual fungi is strongly influenced by the culture medium (Booth, 1971). Morphological characters are the main tool in identifying and describing of a species (Harrington and Rizzo, 1999). This is more useful for quantitative characters, because they can be used in defining species phylogeny (Luckow, 1995). Among the morphological quantitative characters in fungal species, spore size is probably the most commonly character used (Parmasto and Parmasto, 1992). However, if these characters have long been used to identify the pathogenic fungi and to compare the isolates of different origins, analysis has several major drawbacks. These characters are highly variable in many fungi, which limit the scope of their significance in determining population structures. Moreover, in general, these characters (with rare exceptions) cannot be a precise genetic analysis; genes involved in expression are being too numerous (Lourd, 1995).
In this study, we found that P. tritici-repentis isolates showed significant differences in many mprophological characters such as spore size, colony color, etc. grown on the same medium and similar growing conditions. Obtaining fungal isolates information characterized for color, growth, and spore size facilitates further research in the fungus in a multitude of discipline. For example, to study the genetics of a fungus or its interaction with a host, mutants of the fungus are produced. However, to determine if mutagenesis altered these characters, the range of variation in the original isolates for each character must be determined. In addition, isolates with defined characteristics would facilitate studies involving the epidemiology of tan spot.

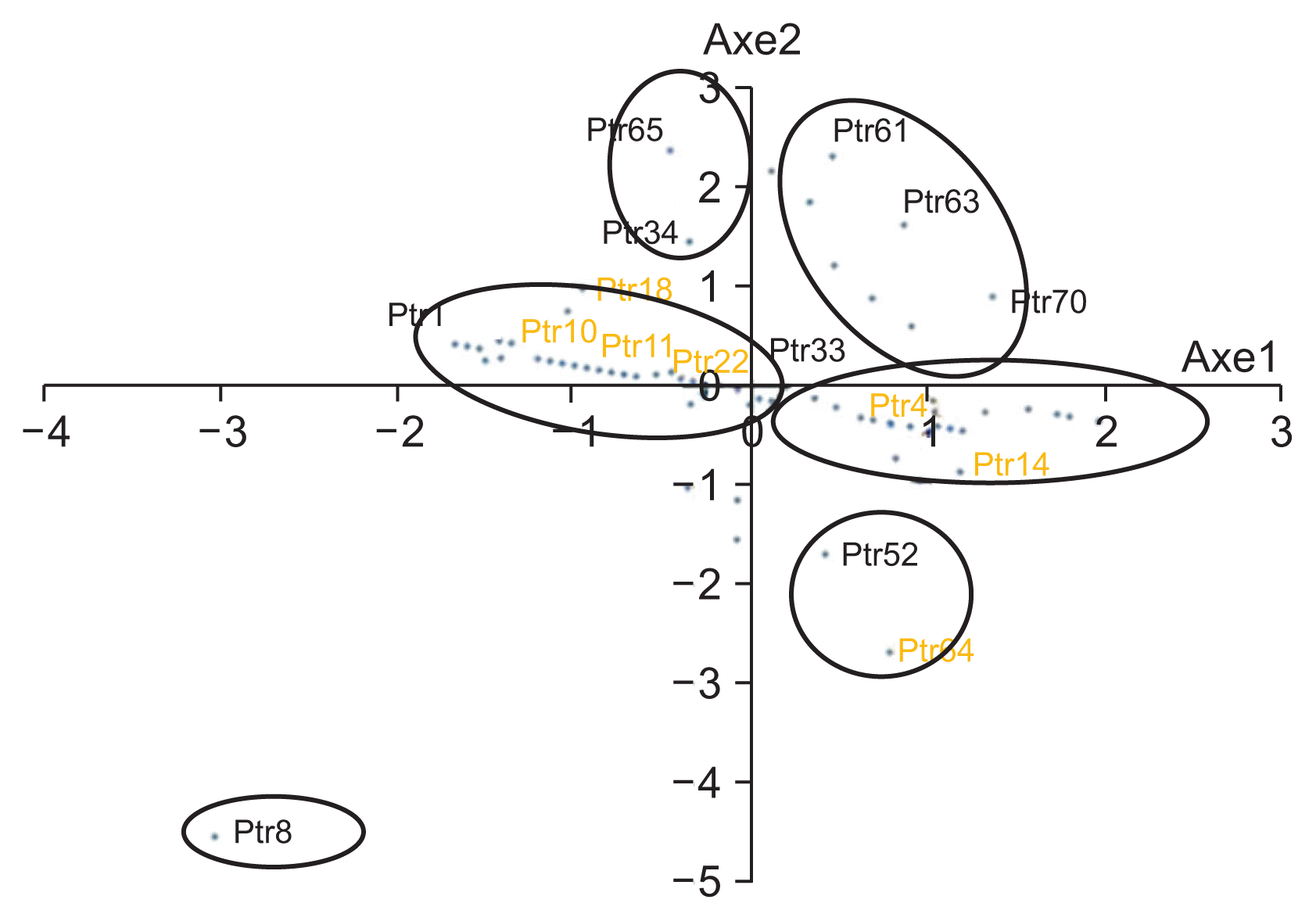
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






