Development and Evaluation of Loop-Mediated Isothermal Amplification Assay for Rapid Detection of Tylenchulus semipenetrans Using DNA Extracted from Soil
Article information
Abstract
Tylenchulus semipenetrans is an important and widespread plant-parasitic nematode of citrus worldwide and can cause citrus slow decline disease leading to significant reduction in tree growth and yield. Rapid and accurate detection of T. semipenetrans in soil is important for the disease forecasting and management. In this study, a loop-mediated isothermal amplification (LAMP) assay was developed to detect T. semipenetrans using DNA extracted from soil. A set of five primers was designed from the internal transcribed spacer region (ITS1) of rDNA, and was highly specific to T. semipenetrans. The LAMP reaction was performed at 63°C for 60 min. The LAMP product was visualized directly in one reaction tube by adding SYBR Green I. The detection limit of the LAMP assay was 10−2 J2/0.5 g of soil, which was 10 times more sensitive than conventional PCR (10−1 J2/0.5 g of soil). Examination of 24 field soil samples revealed that the LAMP assay was applicable to a range of soils infested naturally with T. semipenetrans, and the total assay time was less than 2.5 h. These results indicated that the developed LAMP assay is a simple, rapid, sensitive, specific and accurate technique for detection of T. semipenetrans in field soil, and contributes to the effective management of citrus slow decline disease.
Introduction
The citrus nematode, Tylenchulus semipenetrans Cobb 1913, is an important plant-parasitic nematode that is widely distributed in citrus-growing regions worldwide and causes citrus slow decline disease (El-Borai et al., 2002). T. semipenetrans is a sedentary semiendoparasite, and can infect all Citrus species and most hybrids of citrus in the Rutaceae family (Verdejo-Lucas et al., 2000). Aboveground symptoms of the disease are stunting, slow growth, leaf chlorosis and abscission, reduced fruit size and yield (Verdejo-Lucas and McKenry, 2004). Additionally, the wounds in citrus roots caused by T. semipenetrans are subject to the invasion of plant pathogenic fungi and/or bacteria leading to more severe damage to citrus trees (Duncan, 2009). Yield losses caused by T. semipenetrans are generally 10% to 30% (Verdejo-Lucas and McKenry, 2004). The diagnosis of the disease is very difficult as field symptoms are similar to physiological diseases or other plant diseases. In particular, etiolation symptoms of the disease were often misdiagnosed as citrus Huanglongbing, and these diseased citrus trees were felled or eradicated leading to significant economic losses in some citrus orchards in China (Song et al., 2016). Thus, a rapid and reliable detection method for T. semipenetrans becomes crucially important for effective control of this nematode.
Traditional method for T. semipenetrans identification depends mainly on morphological observation of mature females, males and second-stage juveniles (J2) by microscopy (Inserra et al., 1988; Rashidifard et al., 2015), which is time-consuming, laborious and requires taxonomic expertise. In particular, it is difficult to distinguish J2 of T. semipenetrans from many other plant-parasitic and non-plant-parasitic nematode species microscopically. As an alternative strategy for T. semipenetrans identification, molecular diagnostic methods based on polymerase chain reaction (PCR) have been developed and applied, including PCR-restriction fragment length polymorphism (RFLP) of the internal transcribed spacer region (ITS) (Maafi et al., 2012; Park et al., 2009) and PCR with species-specific primer sets designed from rDNA-ITS (Liu et al., 2011; Maafi et al., 2012). While PCR-based detection methods provided faster, more reliable and more sensitive tools for T. semipenetrans identification when compared with traditional morphological method, they are not generally available for routine practical applications at grassroots quarantine stations and plant protection organizations because expensive and sophisticated laboratory equipment and skilled technicians are needed. Therefore, the development of rapid, simple and cost-effective detection methods is still needed for the specific diagnosis of T. semipenetrans.
Loop-mediated isothermal amplification (LAMP) is a sensitive and rapid nucleic acid amplification technology developed by Notomi et al. (2000) which can amplify DNA under isothermal conditions (60–65°C) in less than an hour. The technique requires Bst DNA polymerase with strand-displacement activity and a set of four to six specially designed primers that recognize six to eight distinct sequences on the target DNA (Nagamine et al., 2002; Notomi et al., 2000). The amplification products are stem-loop DNA structures with several inverted repeats of the target and cauliflower-like structures with multiple loops. LAMP products can be confirmed with the naked eye by adding a fluorescent DNA intercalating dye (e.g., SYBR Green I or propidium iodide) or a metal-ion indicator (e.g., calcein or hydroxynaphthol blue) to the reaction tube and observing the color of the solution (Goto et al., 2009; Hill et al., 2008; Iwamoto et al., 2003; Tomita et al., 2008). All LAMP steps, from amplification to detection, are conducted in one reaction tube, and only a water bath or heating block is required, which make it suitable for both field- and lab-based pathogen detection. To date, LAMP has been successfully developed to detect several plantparasitic nematodes including Bursaphelenchus xylophilus (Kikuchi et al., 2009), Meloidogyne spp. (Niu et al., 2011, 2012), Radopholus similis (Peng et al., 2012) and T. semipenetrans (Lin et al., 2016).
Damage to citrus caused by T. semipenetrans is related to population densities of J2 in the soil (Irshad et al., 2012). Detecting the presence of T. semipenetrans in the soil is essential for both the diagnosis of the disease and many cropping decisions, especially for establishing a citrus nursery site (Liu et al., 2011). Although PCR-based methods described above identified J2 of T. semipenetrans, they were designed to use DNA extracted from isolated individuals that requires a long operation time and experienced personnel (Liu et al., 2011; Maafi et al., 2012; Rashidifard et al., 2015). Hence, a rapid, simple and time-saving nematode extraction method is also needed for quick diagnostic of T. semipenetrans. The development of soil DNA extraction methods has overcome such problems, and these methods have been applied to detect plant-parasitic nematodes in soil, such as Pratylenchus spp. (Yan et al., 2008), Meloidogyne incognita, Heterodera glycines, and Globodera rostochiensis (Min et al., 2012).
Although a LAMP detection method of T. semipenetrans was developed (Lin et al., 2016), the detection sensitivity was relatively lower. The objectives of this study were to develop a rapid, simple, specific and highly sensitive LAMP method for detection of T. semipenetrans directly from soil, and to confirm whether the method was applicable to a wide range of soils naturally infested with T. semipenetrans. In particular, the specificity, sensitivity and field application of the method for detection of T. semipenetrans were assessed.
Materials and Methods
Nematode populations
All nematode populations used in this study were listed in Table 1. T. semipenetrans was isolated from citrus rhizosphere soil in Yongzhou City, Hunan Province, China, and was maintained on a susceptible mandarin orange planted in a plastic pot in a greenhouse. All nematodes had been identified by morphological characteristics and molecular diagnoses.

Species of plant nematode used to evaluate specificity of loop-mediated isothermal amplification (LAMP) assay
DNA extraction
Two methods of DNA extraction were used in this study. Total genomic DNA was extracted from soil using the FastDNA SPIN Kit for Soil (MP Biomedicals, Solon, OH, USA) according to the manufacturer’s instructions, and dissolved to a final volume of 20 μl. DNA of all nematodes in Table 1 was extracted from a single nematode following the method described by Htay et al. (2016), with some modifications. A single nematode was hand-picked using a fine teasing needle and put into a 0.2 ml sterile PCR tube containing 8 μl of distilled water and 1 μl of 10× PCR buffer (Mg2+ free) (Takara, Dalian, China). The tube was frozen in liquid nitrogen for 1 min, then thawed at 95°C in a water bath for 2 min, and the procedure repeated several times. One microliter proteinase K (1 mg/ml) (TransGen Biotech, Beijing, China) was added to the tube and incubated at 56°C for 15 min and consecutively at 95°C for 10 min to inactivate the proteinase K. Finally, the nematode lysis mixture was centrifuged at 12,000g for 1 min and the DNA supernatant was stored at −20°C for future use.
LAMP primer design
The sequence of the 18S-ITS1-5.8S-ITS2-28S rDNA regions from T. semipenetrans (GenBank accession number KX461936) was obtained using universal primers 18s/26s (Vrain et al., 1992), and sequences from Tylenchulus species, including T. semipenetrans (GU433391, FJ969705, and JN112270), T. furcus (JN112284), T. graminis (JN112286), T. musicola (JN112282), and T. palustris (JN112268), were also downloaded from the National Center for Biotechnology Information (NCBI) GenBank database, then aligned and analyzed using BioEdit software (Hall, 1999). Finally, the rDNA-ITS1 sequence was chosen for designing LAMP primers using the Primer Explorer V5 software (http://primerexplorer.jp). Each selected primer sequence is given in Table 2 and their positions are shown in Fig. 1.
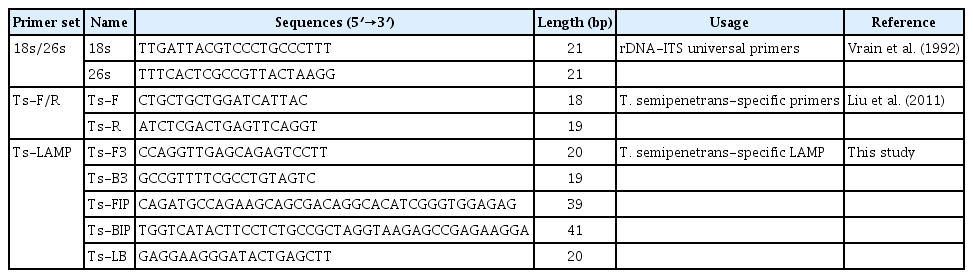
Nucleotide sequences of primers used for conventional PCR and loop-mediated isothermal amplification (LAMP) reaction
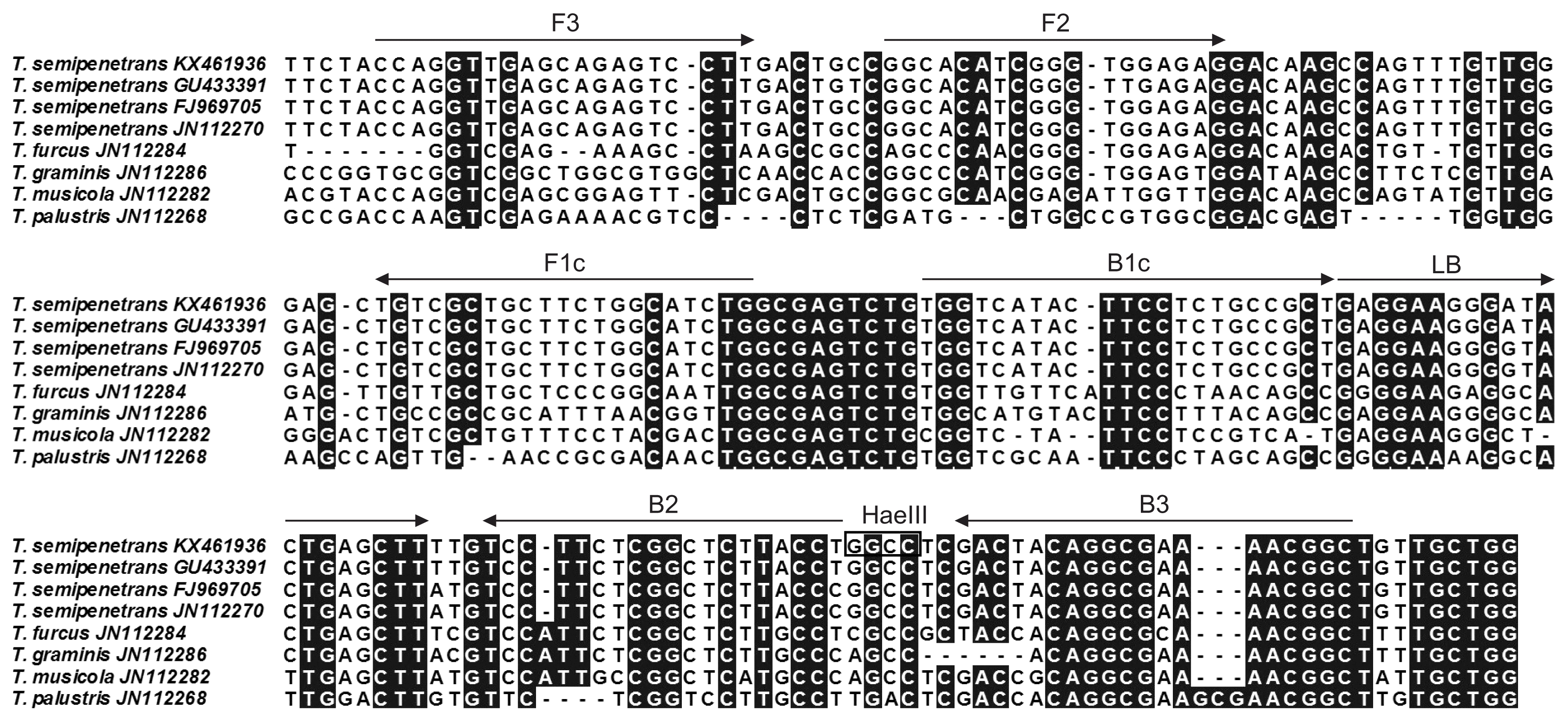
Mutiple alignment of internal transcribed spacer 1 (ITS1) sequences of Tylenchulus semipenetrans and other Tylenchulus species and design for T. semipenetrans loop-mediated isothermal amplification (LAMP) primers. A hyphen indicates a gap. The position of the primers and amplification directions are shown with black arrows. The restriction enzyme site for HaeIII is indicated by small box. The FIP and BIP primers consist of F1c plus F2 and B1c plus B2, respectively.
LAMP reaction
The LAMP reaction was performed according to the method previously described by Notomi et al. (2000) and Nagamine et al. (2002), with some modifications. The optimized LAMP reaction was carried out in a total of 25 μl reaction mixture containing 1.6 μM each of inner primers FIP and BIP, 0.2 μM each of outer primers F3 and B3, 0.4 μM of LB primer, 1.2 mM of dNTPs, 4 mM of MgSO4, 8 U of Bst DNA polymerase (New England Biolabs, Ipswich, MA, USA), 1× ThermoPol reaction buffer (20 mM Tris-HCl, 10 mM KCl, 10 mM (NH4)2SO4, 2 mM MgSO4, 0.1% Triton X-100, pH 8.8) and 1 μl of template DNA. The reaction mixture was incubated in a water bath or heat block at 63°C for 60 min and then heated to 85°C for 5 min to terminate the reaction.
Conventional PCR
The primers 18s/26s were used for obtaining the rDNA-ITS sequence of T. semipenetrans (Vrain et al., 1992). T. semipenetrans-specific primers Ts-F/Ts-R were employed for confirming the specificity of the LAMP assay (Liu et al., 2011). The LAMP outer primers Ts-F3/Ts-B3 was used to verify that the LAMP amplified the correct target and for a sensitivity comparison of LAMP with conventional PCR. PCR amplification reaction was performed in 25 μl reaction mixture containing 2.5 U EasyTaq DNA polymerase (TransGen Biotech), 3 μl of 10× EasyTaq buffer, 0.2 μM each of forward and reverse primers, 0.2 mM of dNTPs and 1 μl of template DNA. The PCR amplification procedure was carried out as follows: initial denaturation at 94°C for 4 min, followed by 35 cycles of denaturation at 94°C for 30 s, annealing (30 s at 52°C for 18s/26s, 1 min at 55°C for Ts-F/Ts-R and 30 s at 60°C for Ts-F3/Ts-B3) and extension at 72°C for 30 s, with a final extension at 72°C for 5 min. The PCR products were separated by 2% agarose gels electrophoresis, stained with ethidium bromide and then photographed under ultraviolet (UV) transilluminator.
Detection of LAMP products
The LAMP amplification results were visually detected with the naked eye or under UV light by adding 2 μl of 1:10 diluted 10,000× concentration fluorescent dye SYBR Green I (Invitrogen, Grand Island, NY, USA) to the mixture and observing the solution’s color. A positive LAMP reaction would turn to green, otherwise, it would remain brown. Additionally, the amplified LAMP products were aslo analyzed by 2% agarose gel electrophoresis, stained with ethidium bromide and then photographed under a UV transilluminator. A positive LAMP reaction would produce the typical ladder-like bands, or else it would produce no DNA bands.
Confirmation of LAMP products
To verify whether the correct target was specifically amplified by LAMP, 186 bp DNA bands were amplified by conventional PCR using the LAMP outer primer pair Ts-F3 and Ts-B3, as previously described. The PCR DNA fragments were extracted from 2% agarose gel using an AxyPrep DNA Gel Extraction Kit (Axygen, Union City, CA, USA). The purified fragments were then cloned into pEASY-T5 Zero Cloning Vector (TransGen Biotech) and transformed into Trans5α Chemically Competent Cells (TransGen Biotech). The recombinant plasmids were extracted from positive clones and sequenced (Sangon Biotech, Shanghai, China).
In addition, LAMP products were digested with HaeIII restriction enzyme (the restriction enzyme site is shown in Fig. 1). The reaction system was performed in a 20 μl volume containing 1 μl of HaeIII (Takara), 2 μl of 10× M Buffer, 6 μl of LAMP products, and 11 μl ddH2O. The reaction was carried out at 37°C for 1 h. The digested products were analyzed on 2% agarose gels electrophoresis, stained with ethidium bromide and then photographed under a UV transilluminator.
Specificity of the LAMP assay
To determine the specificity of the LAMP assay for T. semipenetrans, genomic DNA extracted from 13 nematode species, including one T. semipenetrans, 11 plant-parasitic nematode species in other genera, and one non-parasitic nematode species isolated from citrus rhizosphere soil (Table 1), were assayed by LAMP. Specificity tests were repeated three times.
Sensitivity comparison of LAMP and conventional PCR
Soil was inoculated with T. semipenetrans to evaluate the detection sensitivity of the LAMP assay. The soil (sandy loam) that was not infested with T. semipenetrans was collected from citrus rhizosphere and autoclaved (121°C, 115 kPa) two times for 50 min each time to completely kill all living organisms. One hundred J2 of T. semipenetrans were added to 0.5 g of autoclaved soil using a fine teasing needle under a microscope. Total genomic DNA was extracted directly from inoculated soil using a FastDNA SPIN Kit for Soil (MP Biomedicals) and was serially diluted in ten-fold increments into nine concentrations with sterile distilled water. Different dilutions of genomic DNA were separately detected by LAMP and conventional PCR using the primer pair Ts-F3/B3. Sensitivity tests were repeated three times.
Evaluation of the LAMP assay using natural field soil samples
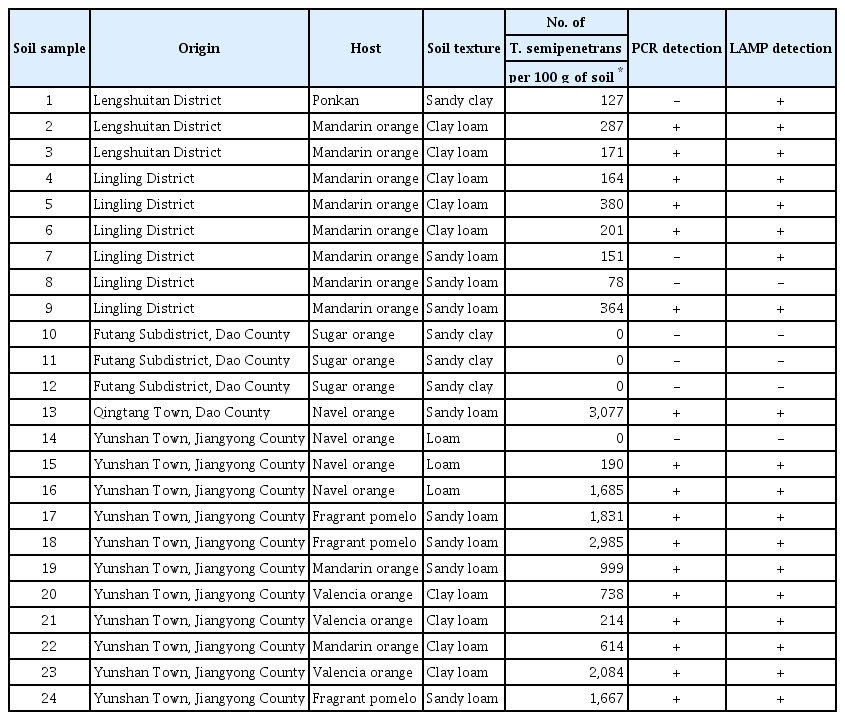
To evaluate the detection capability of the LAMP assay for T. semipenetrans in the field, 24 rhizosphere soil samples were collected from 24 different citrus orchards in Yongzhou City, Hunan Province, China in association with six different citrus crops, and each soil was mixed well. Nematodes were isolated from 100 g of soil using the Whitehead tray method (Whitehead and Hemming, 1965), and the number of T. semipenetrans was counted under a microscope. DNA was extracted directly from 0.5 g of soil using the FastDNA SPIN Kit for Soil (MP Biomedicals) and was separately detected by LAMP and conventional PCR using the primer pair Ts-F/Ts-R, as described previously. Testing of each soil sample was repeated three times.
Results
Detection of LAMP products
Under visual fluorescence detection with SYBR Green I dye, positive reaction containing template DNA of T. semipenetrans turned green, while the negative control retained brown (Fig. 2A). After electrophoresis on agarose gel, the typical ladder-like bands could be observed in the positive sample, whereas the negative control produced no DNA bands (Fig. 2B).
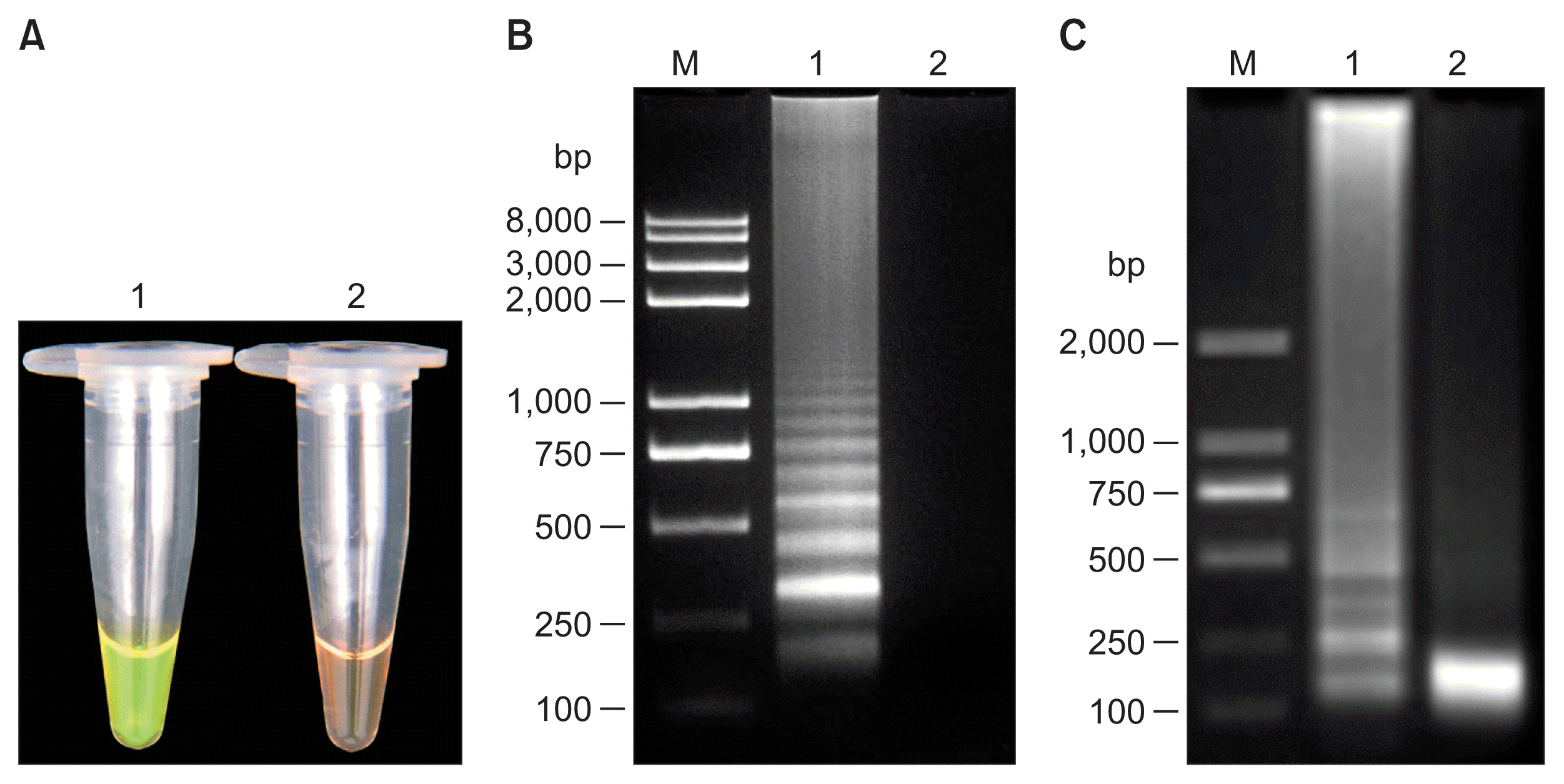
Visual examination, agarose gel electrophoresis and restriction enzyme analysis of loop-mediated isothermal amplification (LAMP) products. (A) LAMP products were detected by adding SYBR Green I fluorescence dye. Tube 1, genomic DNA of Tylenchulus semipenetrans; tube 2, no template control. (B) Agarose gel electrophoresis of LAMP products. Lane M, 8 kb DNA marker; lane 1, genomic DNA of T. semipenetrans; lane 2, no template control. (C) LAMP products were digested by HaeIII. Lane M, 2 kb DNA marker; lane 1, LAMP product without digestion; lane 2, LAMP products digested by HaeIII.
Confirmation of LAMP products
Sequencing results of the recombinant plasmid showed that the 186 bp target sequence was 100% homologous with the rDNAITS1 sequence of T. semipenetrans (KX461936) (data not shown). Moreover, LAMP products were digested completely with HaeIII restriction enzyme (Fig. 2C). The results of the sequence and restriction enzyme analyses confirmed that LAMP products were specifically amplified from the rDNA-ITS1 target sequence of T. semipenetrans.
Specificity of LAMP assay
The specificity of LAMP assay was tested using genomic DNA from T. semipenetrans and 12 other nematode species (Table 3). Only target DNA from T. semipenetrans exhibited a positive reaction, while no positive reaction was observed in the other nematode species (Fig. 3).
Detection of Tylenchulus semipenetrans in natural field soil samples using the Whitehead tray and morphological counting method, PCR and loop-mediated isothermal amplification (LAMP) assay

Specificity assessment of loop-mediated isothermal amplification (LAMP) assay. (A) Visual examination of LAMP products by adding SYBR Green I fluorescence dye. (B) Agarose gel electrophoresis of LAMP products. Tubes and lanes 1–13, genomic DNA of Tylenchulus semipenetrans, Meloidogyne hapla, Pratylenchus coffeae, Helicotylenchus dihystera, Aphelenchus avenae, Filenchus spp., Aphelenchoides spp., Bursaphelenchus xylophilus, Meloidogyne incognita, Cryphodera spp., Tylenchorhynchus spp. Heterodera elachista and Rhabditis, respectively; tube and lane 14, no template control; lane M, 8 kb DNA marker.
Comparison of the sensitivity of LAMP and conventional PCR
The sensitivity test showed that the reaction performed by LAMP using visual fluorescence detection and agarose gel electrophoresis could detect 10−2 J2/0.5 g of soil (Fig. 4A, B). Whereas the reaction carried out using conventional PCR could detect 10−1 J2/0.5 g of soil (Fig. 4C). No amplification was observed in the no template controls.

Sensitivity assessment of loop-mediated isothermal amplification (LAMP) assay and conventional PCR for detection of Tylenchulus semipenetrans. (A) Visual examination of LAMP products by adding SYBR Green I fluorescence dye. (B) Agarose gel electrophoresis of LAMP products. (C) Agarose gel electrophoresis of conventional PCR products. Tubes and lanes 1–9, genomic DNA from 100, 10, 1, 10−1, 10−2, 10−3, 10−4, 10−5, 10−6 J2/0.5 g of soil, respectively; tube and lane 10, no template control; lane M, molecular marker.
Evaluation of the LAMP assay using natural field soil samples
As shown in Table 3, 20 of the 24 soil samples detected T. semipenetrans by the Whitehead tray extraction and microscopic counting method, ranging from 78 to 3,077 nematodes per 100 g of soil. The positive sample detection rate was 17/20 (85.0%) by conventional PCR, and 19/20 (95.0%) by the LAMP assay. The detection limit of conventional PCR and LAMP assay for T. semipenetrans was 164 and 127 nematodes per 100 g of field soil, respectively, as determined by the Whitehead tray extraction and microscopic counting method. No amplification was observed from the non-infected soil samples by either method. All PCR positive samples could be detected by the LAMP assay.
Discussion
T. semipenetrans is the causal agent of citrus slow decline disease, which is a major threat to global citrus production (Duncan, 2009). Rapid and accurate detection of T. semipenetrans in soil is essential for the diagnosis and the effective management of the disease. However, two detection methods currently available have some limitations in routine practical applications. Traditional morphological method is time-consuming and requires specialized knowledge. PCR-based detection methods require expensive instruments and equipment (e.g., thermal cycling). One of the most promising methods for detection of T. semipenetrans is LAMP, which is a simpler operation, more rapid and sensitive reaction, and easier detection compared with conventional PCR. In this study, we developed a simple, rapid and highly sensitive LAMP assay for detection of T. semipenetrans using DNA extracted directly from field citrus rhizosphere soil.
Traditionally, J2 of T. semipenetrans is first extracted from soil using the Baermann funnel method or the centrifugal flotation method before morphological identification or molecular diagnostic. Furthermore, for PCR-based detection methods, DNA extraction is performed using individual nematode from a nematode suspension (Liu et al., 2011; Maafi et al., 2012; Rashidifard et al., 2015). The operational procedures of these methods are time-consuming, and require J2 of T. semipenetrans to be distinguished from the nematode mixture as well, which is not a simple and rapid diagnosis for T. semipenetrans. At present, soil DNA extraction methods have overcome these limitations, and several commercial soil DNA extraction kits have been developed (Ophel-Keller et al., 2008), for example, FastDNA SPIN Kit for Soil (MP Biomedicals) can quickly and efficiently extract PCR-ready genomic DNA directly from soil samples in less than 60 min. In this study, we extracted DNA directly from soil using a commercial kit to research the rapid LAMP assay for T. semipenetrans.
Within the rDNA repeating unit, the ITS regions can be appropriate for nematode species identification (Powers et al., 1997; Ye et al., 2004). In the present study, the LAMP primers were designed for detection of T. semipenetrans based on differences in the rDNA-ITS1 sequences of T. semipenetrans and other Tylenchulus species. The genus Tylenchulus presently contains five species: T. semipenetrans, T. furcus, T. graminis, T. musicola, and T. palustris (Maafi et al., 2012), but except for T. semipenetrans, other Tylenchulus species do not occur in China. To determine the efficiency and specificity of the LAMP assay, we used DNA extracted from T. semipenetrans and 12 other nematode species as templates for the LAMP assay. Only DNA from T. semipenetrans was amplified, and no cross-reaction was found in other nematode species, indicating the LAMP primers designed were specific for detection of T. semipenetrans. Moreover, sequence and restriction enzyme analyses of LAMP products also verified its specificity.
It is generally considered that the sensitivity of LAMP is much higher than that of conventional PCR (Niu et al., 2011, 2012; Peng et al., 2012). Although Lin et al. (2016) reported that the LAMP assay could detect 10−1 J2 of T. semipenetrans, the assay was based on DNA extracted from an individual specimen, not from soil. In our work, the detection limit of conventional PCR using outer primers was 10−1 J2/0.5 g of soil, whereas that of the LAMP assay was 10−2 J2/0.5 g soil. The results showed that the sensitivity of the LAMP assay for detection of T. semipenetrans was 10–100 times higher than conventional PCR method, and previously reported LAMP method by Lin et al. (2016) and PCR method by Liu et al. (2011).
In the field soil sample testing, we collected 24 rhizosphere soil samples from various citrus orchards. It was found that T. semipenetrans existed in 20 of 24 soil samples using the Whitehead tray extraction and microscopic counting method, ranging from 78 to 3,077 nematodes/100 g of soil. The detection limit of the LAMP assay was 127 nematodes/100 g of soil in naturally infested field soil, which was lower than 202 nematodes/100 g of soil by the LAMP method developed by Lin et al. (2016). When the number of T. semipenetrans in soil is very low, 0.5 g of soil for DNA extraction may be too small compared with the Whitehead tray extraction and microscopic counting method that could process 100 g of soil because of the uneven distribution of nematodes in soil. And for this reason, DNA extraction methods from large amounts of soil should be developed for rapid detection of T. semipenetrans in soil. But more importantly, the detection limit of the LAMP assay in natural field soil samples was much lower than the economic threshold level (2,000–4,000 juveniles/100 ml of soil) (Ardakani et al., 2014), indicating that the assay will be useful for disease forecasting and management.
In conclusion, we have established a LAMP assay based on the rDNA-ITS1 sequence of T. semipenetrans and demonstrated that it is a rapid, specific, sensitive and practical method for detection of T. semipenetrans in naturally infested field soil. The LAMP assay can detect T. semipenetrans from soil in less than 2.5 h: 60 min for DNA extraction from soil, 60 min for the LAMP reaction, and 1 min for detection. Therefore, the new LAMP assay could be potentially adaptable for diagnosis and management of citrus slow decline disease caused by T. semipenetrans in the field. Furthermore, this assay lays the foundation for rapid and efficient detection of T. semipenetrans from a large amount of field soil samples.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (31171826), the China Agriculture Research System for Bast and Leaf Fiber Crops (CARS-19-E11), and Hunan Provincial Innovation Foundation for Postgraduate (CX2015B249).