Efficient Transmission and Propagation of Tomato Chlorosis Virus by Simple Single-Leaflet Grafting
Article information
Abstract
Tomato chlorosis virus (ToCV), a member of the genus Crinivirus, has caused an epidemic disease in tomato worldwide. ToCV is phloem-limited and transmitted by whiteflies in a semi-persistent manner, but not by mechanical inoculation. Experimental propagation of ToCV has been performed primarily by using whitefly-mediated inoculation. To develop a simple and convenient method for transmission of ToCV, we investigated grafting single-leaflets from tomato plants infected with ToCV to recipient tomato seedlings. Forty-one of 46 tomato seedlings tested were grafted successfully with single-leaflets infected with ToCV. Among them, 36 seedlings (87.8%) were systemically infected with ToCV and developed typical symptoms. Our results demonstrated that single-leaflet grafting could provide a sufficient amount of inoculum for the transmission of ToCV to the grafted seedlings.
Successful maintenance and propagation of virus strains and isolates are essential for examining their biological, genetic, and evolutionary properties. Although mechanical inoculation is the most commonly used method for propagation of various viruses in laboratories, it may not be employed for all plant viruses because of different modes of action in transmission, particularly for many virus species that are transmitted by vectors (Whitfield et al., 2015). Recent advances in molecular biological techniques have enabled to generate infectious DNA clones of plant virus species and thereby to preserve their genetic and biological characteristics stably (Boyer and Haenni, 1994; Desbiez et al., 2012; Shi et al., 1997). However, the application of these techniques is restricted to certain virus species, limited by genome organization, gene expression strategy and host range.
Tomato chlorosis virus (ToCV), which is a member of the genus Crinivirus in the family Closteroviridae, causes a serious leaf chlorosis disease in tomato plants. In recent years, ToCV epidemics have caused severe economic damage to tomato production worldwide (Tzanetakis et al., 2013). ToCV initially induces interveinal chlorosis symptoms in lower leaves, and then progress towards the upper leaves of the infected plant. The genome of ToCV consists of two segments of positive-sense single-stranded RNAs, RNA1 (8594–8595 nt), and RNA2 (8242–8247 nt) (Wintermantel et al., 2005). The genomic RNA segments are separately encapsidated in flexuous filamentous virus particles varying from 800 to 850 nm in length (Liu et al., 2000). As typical in criniviruses, ToCV is phloem-limited and is not transmissible by mechanical inoculation. In nature, ToCV is transmitted in a semi-persistent manner by phloem-feeding whiteflies belonging to two genera, Bemisia (B. tabaci) and Trialeurodes (T. vaporariorum and T. abutiloneus) (Wintermantel and Wisler, 2006). Therefore, the maintenance and propagation of ToCV isolates are usually performed by whitefly-mediated inoculation for experimental uses. However, individual inoculation with whiteflies is labor-intensive and complicated and requires the maintenance of whitefly colonies in addition to the development of a controlled inoculation protocol. Here, we developed a simple and efficient method for ToCV transmission by grafting with single leaflets from infected tomato plants as sources of inocula. To compare the efficiency of ToCV transmission by grafting, the same method was tested for the transmission of tomato yellow leaf curl virus (TYLCV) and for co-transmission of both viruses in parallel.
The viral sources used in this study were the ToCV isolate IKC and the TYLCV isolate IKY. Both isolates were obtained from the naturally infected tomato plants collected from a commercial field in Korea and maintained in tomato cv. Tenten by periodic transmission with B. tabaci.
Six-week-old tomato seedlings (cv. Tenten) were used as recipient plants for single-leaflet grafting inoculation (Fig. 1A). Leaflets for grafting inoculation were taken from ToCV- or TYLCV-infected tomato plants designated as source plants for viral inocula. Prior to grafting inoculation, the absence and presence of the viruses in all recipient and source plants, respectively, were confirmed by PCR (for TYLCV) and RT-PCR (for ToCV) using specific primer pairs (For TYLCV, 5′-GTCAACCAATCAAATTGCATCCTCAA-3′ and 5′-GTCCAAAATCCATTGGGC-3′; For ToCV, 5′-CTACACGCTTACDCCTTATT-3′ and 5′-ACACCGATTCATTTCAACAT-3′). For grafting, a small incision was made on the stem of the recipient seedling by removing the first leaf from the node (Fig. 1B). The beveled cutting tip of the petiole of the inoculum leaflet was grafted onto the incision and the tip of the grafted leaflet was tied to the stem of the recipient seedling with a thin strip of Parafilm (American Can Co., Greenwich, CT, USA) in order to keep the leaflet upright while the tissues healed (Fig. 1C). The newly grafted tomato seedlings were kept under the shade in an air-conditioned, insect-free greenhouse maintained at 26°C during daytime and 20°C at night. In addition, the grafted leaflets were misted with water several times daily to avoid wilting of the leaflets. The grafted plants were maintained under these conditions for two weeks after grafting. Grafting leaflets from virus-free healthy tomato plants was included as a negative control (mock). Four independent sets of the grafting inoculation experiment were performed.

Grafting of single leaflets to tomato seedlings. (A) Six-week-old tomato seedlings (cv. Tenten) were used as recipient plants for single-leaflet grafting inoculation. (B) The first leaf was removed from the stem of the recipient seedling to create an incision for grafting. (C) Leaflets were taken from tomato chlorosis virus- or tomato yellow leaf curl virus-infected tomato plants. The beveled cutting tip of the petiole of the leaflet was grafted onto the incision and tied to the stem with a thin strip of Parafilm. (D) A representative tomato leaflet grafted successfully to the stem of the recipient tomato plant. (E–H) Enlarged images of the areas indicated by boxes in A–D, respectively.
At four weeks after grafting, the grafted tissues were checked. In 162 out of the 184 cases (88%), the grafted tissues had healed successfully in place (Fig. 1D). Five weeks after grafting, nucleic acids were extracted from the upper systemic leaves of the recipient tomato plants that were grafted successfully and subjected to RT-PCR and PCR using the specific primer pairs mentioned above to detect ToCV and ToCV, respectively. The representative gel images of RT-PCR and PCR detection of ToCV and TYLCV, respectively, are shown in Fig. 2. The results of the virus transmission are summarized in Table 1. For each inoculum, a total of 46 tomato seedlings were tested in four independent experiments. In the case of grafting inoculation with ToCV-infected leaflets, 41 plants were grafted successfully and, among them, 36 plants (87.8%) were infected with ToCV. Among the 40 plants grafted successfully with TYLCV-infected leaflets, 36 plants (90.0%) were infected with TYLCV. To examine whether the co-infection of ToCV and TYLCV affects the transmission efficiency of each other upon graft inoculation, 45 tomato seedlings were inoculated by grafting with the leaflets co-infected with ToCV and TYLCV. Interestingly, 35 out of the 39 plant that were grafted successfully were infected with both viruses. In addition, there was no case of single infection, suggesting that the transmission efficiencies of ToCV and TYLCV upon graft inoculation are not suppressed by co-infection. Collectively, our results demonstrated that graft inoculation with virus-infected leaflets is a highly efficient method of transmitting two different types of viruses that are not transmissible by mechanical inoculation in tomato. The grafted tomato plants were observed over a period of two months after inoculation (Fig. 3).
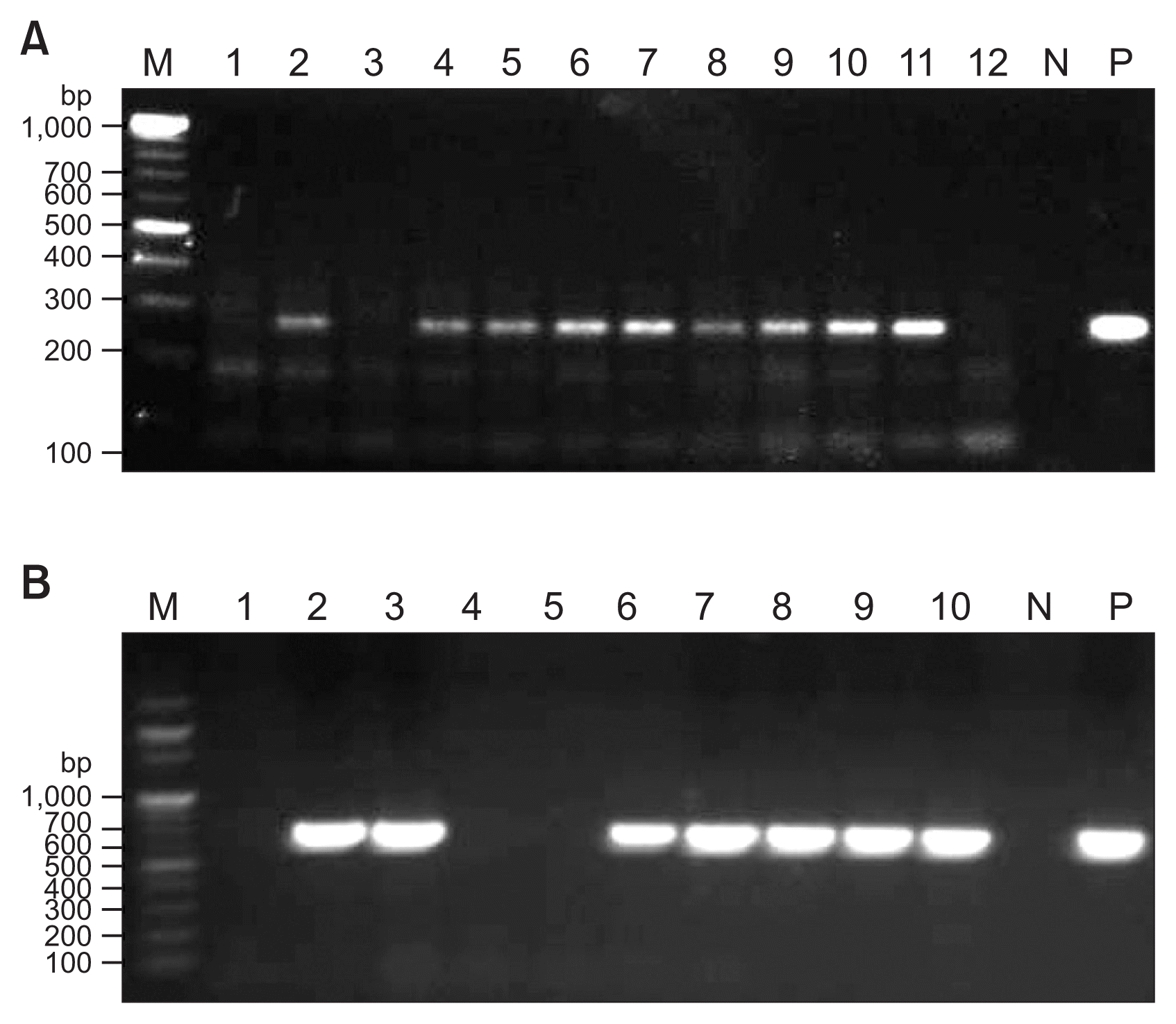
Molecular detection of tomato chlorosis virus (A) and tomato yellow leaf curl virus (B) by RT-PCR and PCR, respectively, from the upper systemic leaves of the single leaflet-grafted tomato plants. PCR amplicons were visualized under ultraviolet light in 1% agarose gel (0.5× TBE buffer) containing 1 μg/ml ethidium bromide. M, 100 bp DNA ladder (Enzynomics, Daejeon, Korea); N, negative controls of PCR reactions; P, positive controls of PCR reactions.

Incidence of infection in tomato seedlings grafted with single-leaflets infected with ToCV, TYLCV, or ToCV + TYLCV

Representative images of the tomato plants grafted with either tomato chlorosis virus (ToCV), tomato yellow leaf curl virus (TYLCV), or ToCV + TYLCV at two months after grafting. No obvious symptoms were observed in the tomato plants grafted with ToCV (B) when compared with untreated healthy tomato plants (A). The tomato plants grafted with TYLCV (C) or ToCV + TYLCV (D) developed typical symptoms of TYLCV. Infection of the grafted tomato plants with ToCV and/or TYLCV was confirmed by RT-PCR and PCR using specific primers.
To examine whether serial passages of ToCV by grafting inoculation can result in accumulation of mutations in the viral genome, the partial genomic sequence of the ToCV isolate IKC RNA2 was analyzed after four serial passages in tomato over 6 months. Before beginning the first grafting, nucleic acids were extracted from the initial source tomato plant infected with ToCV-IKC and subjected to analyze the 3′-half nucleotide sequence of ToCV RNA2 covering ORF6 to ORF9 (approximate nucleotide positions 4355–8027) by amplifying the region using a primer pair (5′-ACACTGGTGATAACGGTGGTG-3′ and 5′-AACGAACTATTGGAGGTTGTGAGT-3′). The sequence of the amplified product was determined by direct sequencing. After four serial passages, nucleic acids were extracted from systemic leaves of the grafted plant and subjected to analyze the 3′-half nucleotide sequence of RNA2 of ToCV progenies as described above. Comparison of two sequences showed that three nucleotide mutations were introduced during four passages: one nucleotide mutation in ORF7 (from T to G at nucleotide position 7099) and two mutations in ORF9 (from G to A at nucleotide position 7747 and from A to G at nucleotide position 7764). The mutation rates of RNA viruses may vary and depend on various biological and environmental constraints. It was suggested that the rates of spontaneous mutations for some RNA viruses are in the range of 10−4 and 10−6 mutations per nucleotide and generation (Drake and Holland, 1999; Tromas and Elena, 2010). Our results showed that single-leaflet grafting is sufficient for transmission of a viral progeny population containing mutations.
Vegetative grafting has been used for inoculation and propagation of many systemic plant pathogens belonging to viruses or bacteria (Damsteegt et al., 2007; Hilf and Lewis, 2016; Kasrawi et al., 1988; Nie and Singh, 2001). One of the advantages of graft inoculation is that it allows the continuous exposure of a recipient plant to a high level of inoculum. Once the vascular system is connected between the source and recipient tissues, pathogens can move from the inoculum and infect the recipient plant. Thus, grafting has been applied for the inoculation of various viral pathogens that are not easily transmissible by mechanical inoculation. In this study, we demonstrated that a single-leaflet from a tomato plant infected with ToCV and/or TYLCV can be successfully grafted to a tomato seedling and provide sufficient inocula for the transmission of the viruses to the grafted seedlings. Thus, this single-leaflet grafting procedure can provide increased experimental flexibility for studying various biological aspects of the ToCV-tomato pathosystem, including the screening of resistant cultivars, maintenance of virus isolates, and examination of virus evolutionary properties.
Acknowledgments
This research was supported in part by grants from the Agenda Program (PJ0120132017) funded by the Rural Development Administration of Korea and the Intramural Support Program (1403-20160062) funded by Seoul National University.