

Introduction
Land plants absorb nutrients and water from soils. Seeing at an evolutionary point of view, mutualism between ancient plants and diverse microorganisms, especially endomycorrhizae, led the most important leap so that plants successfully colonized in the land (Pirozynski and Malloch, 1975; Parniske, 2008). Plant-growth promoting rhizobacteria (PGPRs), that colonize root surface and rhizosphere, positively regulate plant growth, and also enhance disease resistance against plant’s natural enemies (Kloepper, 1994; Lugtenberg and Kamilova, 2009). The beneficial effects of PGPRs are mainly due to production of various metabolites that can directly and indirectly modulate plant physiology related to plant growth and immunity (Kloepper and Schroth, 1981; Lugtenberg and Kamilova, 2009; Zamioudis and Pieterse, 2012).
Induced-resistance is often classified into two groups based on kinds of causal biological agents and plant signaling pathways triggered by the inducers. First, induced-resistance accelerated by PGPRs usually harnesses jasmonate (JA)-mediated signaling machinery and the induced-resistance is called as induced systemic resistance (ISR) (Pieterse et al., 1998; Choudhary et al., 2007). On the other hand, foliar immunization with virulent and/or avirulent pathogens, which induce severe necrosis and hypersensitive response, respectively, also activates systemic acquired resistance (SAR) (Ross, 1961; Ryals et al., 1996). In case of SAR, plants need salicylic acid (SA) accumulation both in local and distal systemic tissues, and generate long distance mobile signals to establish successful immune response against secondary pathogen infection (Malamy et al., 1990; Gaffney et al., 1993; Dempsey and Klessing, 2012). Several genetic analyses reveal that mutant plants of Arabidopsis (A. thaliana L.) defective in JA- and SA-dependent signaling are insensitive to exogenous applications of PGPRs and SAR-inducing microbes (Pieterse et al., 1998; Jung et al., 2009; Zamioudis and Pieterse, 2012).
The exogenous stimulation of plant immune response by microorganisms can directly turn on SA- and/or JA-dependent defense responses regardless of subsequent pathogen infection (Hahn, 1996; Pieterse et al., 1998; Wiesel et al., 2014). Even if the treatment effectively confers disease resistance, a proper energy allocation is disturbed, which results in growth defect in plants (Walters et al., 2013). A different mode of action of beneficial microorganisms is to prime plants to rapidly and strongly activate defense-related events during pathogen infection (Conrath et al., 2002; Conrath, 2011). In this case, plants do not show any significant physiological changes without pathogen infection (Conrath et al., 2015). Thus the priming of defense response is noteworthy phenomenon for effective management of plant diseases (van Hulten et al., 2006).
Bacillus is one of well-identified genera, acting as beneficial PGPRs (Kloepper and Ryu, 2006; Choudhary and Johri, 2009). In addition to direct formulation of Bacillus spp. as PGPR inoculants, tens of different antibiotics were identified in the genus (Compant et al., 2005). During the last 20 years, we isolated B. vallismortis EXTN-1, B. amyloliquefaciens and B. vallismortis BS07 strains from the rhizosphere soils of chili pepper (Capsicum annuum L.), that not only promote plant growth, but also control the plant disease (Park et al., 2001, 2006a, 2006b, 2007, 2013). Especially, treatment with the B. vallismortis BS07 on pepper seedlings confers disease resistance against infection with Phytophthora capsici and Colletotrichum acutatum, the most destructive pathogens to pepper plants (Park et al., 2013). Additionally, PATHOGENESIS-RELATED PROTEIN1 (PR1, a marker for SA-dependent defense response), but not PDF1.2 (a marker for JA-dependent defense response), is expressed in leaves of Arabidopsis 24 hours after treatment with the B. vallismortis BS07 strain. Interestingly, PR1 expression in B. vallismortis BS07-treated plants disappeared in leaves of Arabidopsis NahG plants expressing salicylate hydroxylase that converts SA to catechol (Park et al., 2013). These results suggest that induced-resistance by the B. vallismortis BS07 strain may require the accumulation of SA in plants.
Cyclic dipeptides (CDPs) are one of the simplest compounds that are derived from bacteria to human (Prasad, 1995; Bellezza et al., 2014). Bacillus genus also generates lots of CDPs conjugated with lipid molecules, such as iturins, fengycin, and surfactins, which trigger activation of plant immune response (Ongena and Jacques, 2008; Park et al., 2016). In addition, CDPs produced by Streptomyces sp. and Bacillus spp. have antibacterial and antifungal activities (Kumar et al., 2014; Wattana-Amorn et al., 2016). These previous reports strongly suggest that CDPs are good candidates for plant disease management.
Here we introduce novel biological activities of four different CDPs to induce disease resistance in Arabidopsis plants. Both a synthetic CDP, cyclo(Gly-L-Pro) and three different CDPs, cyclo(L-Ala-L-Ile), cyclo(L-Ala-LLeu), and cyclo(L-Leu-L-Pro) isolated from B. vallismortis BS07, which do not have direct antimicrobial activity in vitro, require certain proteins important for SA- and/or JA-dependent defense responses to successfully inhibit bacteria growth in Arabidopsis. Additionally, exogenous treatment of these CDPs can prime plants to strongly express PR4 gene involved in JA-related defense in Arabidopsis.
Materials and Methods
Plants
Wild-type Arabidopsis thaliana L. ecotype Columbia-0 (Col-0) and mutant plants, whose parental ecotype is an ecotype Col-0, grew in an environmentally controlled growth chamber (21±1°C, 50-60% relative humidity, 12 h day and 12 h night). phytoalexin deficient4 (pad4), salicylic acid induction deficient2 (sid2), nonexpressor of pathogenesis-related proteins1 (npr1), ethylene receptor1 (etr1), jasmonate response1 (jar1) and jasmonate insensitive1 (jin1) mutants were used in this study (Bleecker et al., 1988; Staswick et al., 1992; Berger et al., 1996; Cao et al., 1997; Jirage et al., 1999; Nawrath and Métraux, 1999). Seeds of wild type and mutant plants were sterilized with 50% bleach solution, and kept at 4°C for 3 days until sowing. Plants were watered twice a day until experiments.
Extraction and synthesis of CDPs
Tryptic soy agar media (20 L) (15 g tryptone, 5 g soytone, and 5 g NaCl/L) incubated with B. vallismortis BS07 at 28°C for 7 days were harvested with methanol (20 L × 2) twice and the extracts were concentrated under vacuum. The concentrate was subsequently dissolved in 50% aqueous methanol (500 mL) followed by extractions with n-butanol (4 L × 3). The n-butanol phase was evaporated to dryness, affording 43.5 g of a brown gum, which was chromatographed on a C18 flash column (90 id × 70 mm, C18, 40 × 63 μm, Merck) using a 10% stepwise gradient elution of increasing methanol concentration in H2O (1 L each) to afford eleven fractions (A to K: 28.0 g, 3.0 g, 2.0 g, 3.0 g, 2.0 g, 3.0 g, 1.5 g, 4.5 g, 2.0 g, 1.5 g, and 1.0 g, respectively). Fractions C (2.0 g) was fractionated on a normal phase MPLC (50 id × 220 mm, Silica Redi Sef Rf: Teledyne Isco) eluted with ethyl acetate/methanol/H2O (80:20:0 to 30:63:7) to give eighteen sub-fractions (C1 to C18) from fraction C. Sub-fraction C2 (130 mg) was purified by preparative C18 HPLC (Cosmosil, 5C22-AR-II, 20 id × 250 mm, 25-30% aqueous methanol) to [6.0 mg, tR 25.0 min, cyclo(LAla-L-Ile)], [12.0 mg, tR 28.0 min, cyclo(L-Ala-L-Leu)], and [2.5 mg, tR 39.0 min, cyclo(L-Leu-L-Pro)]. The cyclic dipeptides were synthesized according to the previously reported method (Thajudin et al., 2010). Briefly, N-Boc protected amino acid (1 mmol) was dissolved in dimethyl formamide. N,N-diisopropylethylamine (1.1 mmol) and 2-(1H-benzotriazol-1-yl)-1,1,3,3,-tetramethyluronium hexa-fluorophosphate (1.1 mmol) were added at room temperature, followed by addition of amino acid methyl ester HCl salt (1 mmol). The mixture was stirred for 3 hours. The reaction mixture was diluted with ethyl acetate, washed with water, 1 M NH4Cl solution, saturated sodium bicarbonate solution, and saline. The resulting linear dipeptide was dissolved in water and autoclaved at 130°C for 4 hours and water was evaporated under vacuum. The remaining residue was purified by silica gel column chromatography (hexane/ethyl acetate/MeOH or CH2Cl2/MeOH).
Chemical treatment
Different concentrations of cyclo(Gly-L-Pro), cyclo(L-Ala-L-Ile), cyclo(L-Ala-LLeu), and cyclo(L-Leu-L-Pro) were dissolved in distilled water before treatment. Both 1 and 10 ppm (part per million) of each CDP solution were infiltrated in leaves of Arabidopsis with a needless syringe and 100 ppm of them was applied onto plants with a sprayer 1 day prior to pathogen infection. The CDP-treated plants were cultivated in a typical growth conditions as described above.
Bacterial inoculation
A virulent Pseudomonas syringae pv. maculicola ES4326 was employed in this study. The strain was cultured in King’s B liquid media (10 g proteose peptone, 1.5 g K2HPO4, 15 g glycerol and 5 mM MgSO4/L) supplemented with 50 μg/ml streptomycin at 28°C. The freshly cultured strain was diluted to OD600=0.0001 (to test disease resistance) or OD600=0.01 (to test gene expression) in 10 mM MgSO4. We infiltrated the diluted bacteria into leaves of wild type and mutant plants with a needleless syringe. After infection, the infected plants were covered with a transparent plastic dome to maintain high humidity, and incubated in a growth chamber during 3 days (21±1°C, 12 h day and 12 h night). Number of bacteria was counted on day 3 after infection.
Induction of priming response and quantitative real-time PCR
One ppm of these CDP solutions were infiltrated into leaves of wild-type Arabidopsis. One day after treatment, P. syringae pv. maculicola ES4326 (OD600=0.01) was infiltrated into the pretreated leaves with each CDP. During infection, plants were kept in a growth chamber. The infiltrated leaves were collected at the indicated time after infection. Total RNAs were isolated with the TriZol® reagent according to the manufacturer’s instruction (Thermo Fisher Scientific, MA, USA). To minimize DNA contamination, TURBO DNase (Thermo Fisher Scientific) was treated in the total RNAs at 37°C for 30 min. About 1 μg of total RNA was used to synthesize first-strand cDNAs with Superscript II reverse transcriptase (Thermo Fisher Scientific). qRT-PCR was performed with SYBR Green PCR mixture (Takara Bio, Japan) using the cycling program as following: 95°C for 5 min. followed by 40 cycles at 95°C for 15s, 60°C for 15s and 72°C for 35s (Bio-Rad, CA, USA) (Jung et al., 2009; Lee et al., 2016). Actin2 (At3g18780) was used as the internal control. Oligo-nucleotide sequences used in this study will be provided under a request.
Results and Discussion
Exogenous treatment with CDPs effectively inhibited bacterial growth in Arabidopsis
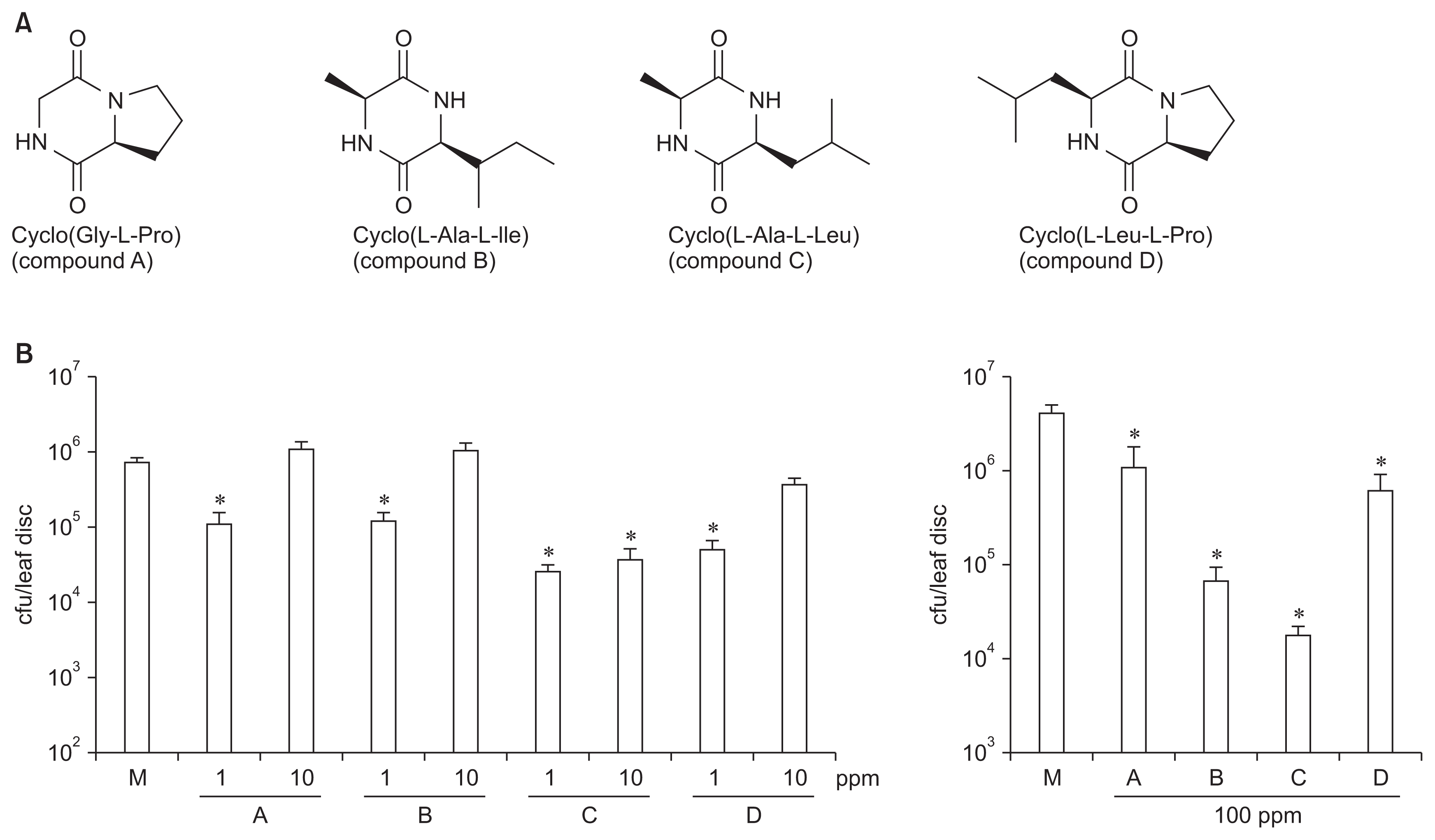
In order to examine whether or not CDPs could inhibit microbial growth in Arabidopsis, first, we synthesized cyclo(Gly-L-Pro) (compound A), that is one of well-documented CDPs from Aspergillus spp, according to the literature (Thajudin et al., 2010; Ortiz-Castro et al., 2011; Li et al., 2012). Additionally, we also recently isolated 17 different CDPs from B. vallismortis BS07 strain. By means of initial screening to identify bioactive compounds, we selected three different CDPs, cyclo(L-Ala-L-Ile) (compound B), cyclo(L-Ala-LLeu) (compound C), and cyclo(L-Leu-L-Pro) (compound D) conferring disease resistance on several crop plants (data not shown). The molecular structures of these CDPs was described in Fig. 1A.
In order to characterize function of these CDP in plants, we used Arabidopsis plants. Wild-type Arabidopsis Col-0 plants were cultivated in potting mixture (Dongbu Farm Hannong, Korea), in an environmentally controlled growth chamber. Different concentrations of each CDP (1, 10, and 100 ppm) were prepared in distilled water. Each solution was treated on plants 1 day prior to inoculation, as mentioned above. A virulent P. syringae pv. maculicola ES4326 strain was infiltrated in the pretreated leaves with the each compound (OD600=0.0001). Treatment of 1 ppm of the compounds inhibited Pseudomonas growth in plants, as compared with mock-treatment (Fig. 1B, left). Direct infiltration of 10 ppm of the compounds, however, was not effective, except compound C (Fig. 1B, left), indicating that concentration of the CDPs may be important for controlling bacterial disease. Additionally, number of bacteria was also reduced in plants applied with 100 ppm of each CDP (Fig. 1B, right). Next, to test whether or not these CDPs had a direct antimicrobial activity to phytopathogenic fungi, and oomycetes, we performed a paper disc-agar diffusion assay of the compound. Alternaria alternata, C, acutatum, Fusarium oxysporum, Rhizoctonia solani, and P. capsici were co-incubated with paper discs containing different concentration of the compound. Mycelial growth of these microorganisms was not inhibited by these compounds in vitro (data not shown), which indicate that these CDPs do not have direct antifungal and antioomycete activity. Thus, we hypothesize that biological activity of these CDPs is due to the activation of plant’s built-in defense response, rather than direct inhibition of microbial growth in plants.
Induced-resistance triggered by CDPs required several key regulators important for SA- and/or JA-dependent defenses
As described above, it is known that disease resistance induced by PGPRs usually depends on JA-related defense response (Pieterse et al., 1998; Zamioudis and Pieterse, 2012). On the other hand, systemic resistance initiated by primary local infection with avirulent or virulent pathogens relies on SA-related events both in local and distal tissues and generation of long distance mobile signals in local tissues (Dempsey and Klessing, 2012). However, it is not yet clear which defense-related signaling pathways are activated by these CDPs. To address this question, first we prepared 100 ppm of each CDP and applied them on various Arabidopsis mutants lacking a SA-dependent event as following; pad4 (SA regulation) (Jirage et al., 1999), sid2 (SA biosynthesis) (Nawrath and Métraux, 1999), and npr1 (SA responsiveness) (Cao et al., 1997). Twenty six-day old plants were used for the experiments. One day after treatment, a virulent P. syringae pv. maculicola ES4326 strain (OD600=0.0001) was infiltrated into leaves of plants, and number of bacteria was counted in the infected leaves 3 day after inoculation. Bacterial growth was only reduced in sid2 mutant, but not pad4 and npr1 mutants, as compared with that in mock-treated plants, after treatment of compound A, which indicates that compound A requires PAD4 and NPR1 for inducing disease resistance (Fig. 2A). In case of compound B, PAD4, SID2, and NPR1 are inessential to induce resistance response against Pseudomonas infection, because these mutant plants were still sensitive to application of compound B (Fig. 2A). Exogenous treatment of compound C effectively inhibited bacterial growth in leaves of pad4 mutant, but not sid2 and npr1 (Fig. 2A). The result proposes that the compound C requires SA biosynthesis and responsiveness for establishing disease resistance in Arabidopsis. On the other hand, mutation of SID2 and NPR1 did not affect compound D-induced resistance in plants, whereas PAD4 was necessary for the resistance (Fig. 2A). Based on these results, we conclude that each CDP tested in this study can differentially influence SA-dependent defense response in Arabidopsis.
Exogenous treatment of B. vallismortis strain BS07 induces PR1 expression, but not PDF1.2, in Arabidopsis (Park et al., 2013). In general, however, JA-dependent signaling is important for induced-resistance triggered by PGPRs. Since these CDPs were produced by Aspergillus sp. (compound A), and B. vallismortis (compound B, C, and D), we expected that key components of JA-dependent events might also be required for the resistance response induced by these CDPs. To test this, we applied 100 ppm of these CDPs on etr1 (ethylene perception) (Bleecker et al., 1988), jar1 (JA-Ile production) (Staswick et al., 1992), and jin1 (JA responsiveness) (Berger et al., 1996) mutants 1 day prior to P. syringae pv. maculicola ES4326 inoculation (OD600=0.0001). Treatment of both compound A and B failed to suppress bacterial growth in etr1 and jin1 mutants, whereas bacterial growth was decreased in leaves of jar1 mutants, as compared with those in mock-treated plants (Fig. 2B). These indicate that both compound A and B require ETR1 or ETR1-regulated proteins and JIN1 to trigger disease resistance against Pseudomonas infection. In case of compound C and D, numbers of bacteria in etr1, jar1, and jin1 mutants were comparable with those in mock-treated plants, which proposes that ethylene and JA-related events are crucial for compound C- and/or D-induced disease resistance in Arabidopsis.
PR4 transcription strongly occurred in the CDP-treated plants at early infection phase
There are two different mechanisms of action of exogenous stimuli to execute induced-resistance in plants. Benzothiadiazole (BTH), a well-known elicitor for plant disease resistance, directly activates SA-related defense response after treatment (Lawton et al., 1996). Priming of defense response is another mechanism of action for induced-resistance (Conrath et al., 2002). Exogenous treatment of chemical primers does not cause any detectable dramatic shift of defense response before pathogen infection. However, immune response is strongly activated in primed plants more than in non-primed plants (Jung et al., 2009; Conrath et al., 2015). Indeed, expression levels of a few defense-related genes in Arabidopsis treated with compound A, B, C, and D were similar to those in mock-treated plants in the absence of pathogen infection (data not shown). The observation allowed us to examine whether or not these chemical inducer acted as primers for establishing disease resistance response in Arabidopsis. To test the possibility, 100 ppm of each CDP was applied on wild-type Arabidopsis. At 24 h after treatment, P. syringae pv. maculicola ES4326 (OD600=0.01) were infiltrated into leaves of CDP-treated plants, and the infected leaves were taken 12 h and 18 h after inoculation (Fig. 3A). To test whether or not the treatment primed plants to rapidly express defense-related genes, we examine mRNA expression of PR1 (At2g14610) and PR4 (At3g04720, Hevein-like protein, a marker for JA-dependent defense response) after subsequent pathogen infection (Fig. 3B) (De Vos et al., 2005; Lorenzo and Solano, 2005). Expression of PR1 in compound A, B, and D-treated plants were comparable with that in mock-treated plants after Pseudomonas infection, whereas PR1 expression level in compound C-treated plants was higher than mock-treated plants 18 hours after infection. This strongly suggests that compound C may have an ability to prime SA-dependent defense responses in Arabidopsis after pathogen infection (Fig. 3B, left panel). Interestingly, exogenous treatment of these CDPs triggered strong mRNA expression of PR4 in Arabidopsis leaves 12 h after subsequent pathogen infection (Fig. 3B, right panel). However the expression did not strongly continue until 18 h after infection in the CDP-treated plants, as compared with that in mock-treated plants. These results strongly suggest that CDPs tested in this study mainly trigger expression of JA-responsive genes during pathogen infection in Arabidopsis.
In conclusion, CDPs from B. vallismortis BS07 strain had different mode of action to induce disease resistance in Arabidopsis. Especially, the facts that most of compounds require JA-responsiveness and enhanced PR4 expression at early infection phase demonstrate that the CDPs tested in this study can prime JA-dependent defense response during pathogen infection. How do plants recognize these CDPs from soil-born bacteria? We are confident that the perception mechanism will be elucidated using genetic analysis in the near future.

 ), B (
), B (
 ), C (□), D (▧) and 1 μM of benzothiadiazole (BTH) (▨) were applied in leaves of wild-type Arabidopsis plants 1 day prior to inoculation. Inoculated leaves were taken for total RNA extraction at the indicated time points after infection with Pseudomonas syringae pv. maculicola ES4326 (OD600=0.01). (B) mRNA expression of PR1 and PR4 genes in the leaves of wild-type Arabidopsis after infection. An Arabidopsis Actin2 gene (At3g18780) was used as an internal reference gene. Expression levels of PR1 and PR4 were calculated by the comparative CT method (2−ΔΔCT) (
), C (□), D (▧) and 1 μM of benzothiadiazole (BTH) (▨) were applied in leaves of wild-type Arabidopsis plants 1 day prior to inoculation. Inoculated leaves were taken for total RNA extraction at the indicated time points after infection with Pseudomonas syringae pv. maculicola ES4326 (OD600=0.01). (B) mRNA expression of PR1 and PR4 genes in the leaves of wild-type Arabidopsis after infection. An Arabidopsis Actin2 gene (At3g18780) was used as an internal reference gene. Expression levels of PR1 and PR4 were calculated by the comparative CT method (2−ΔΔCT) (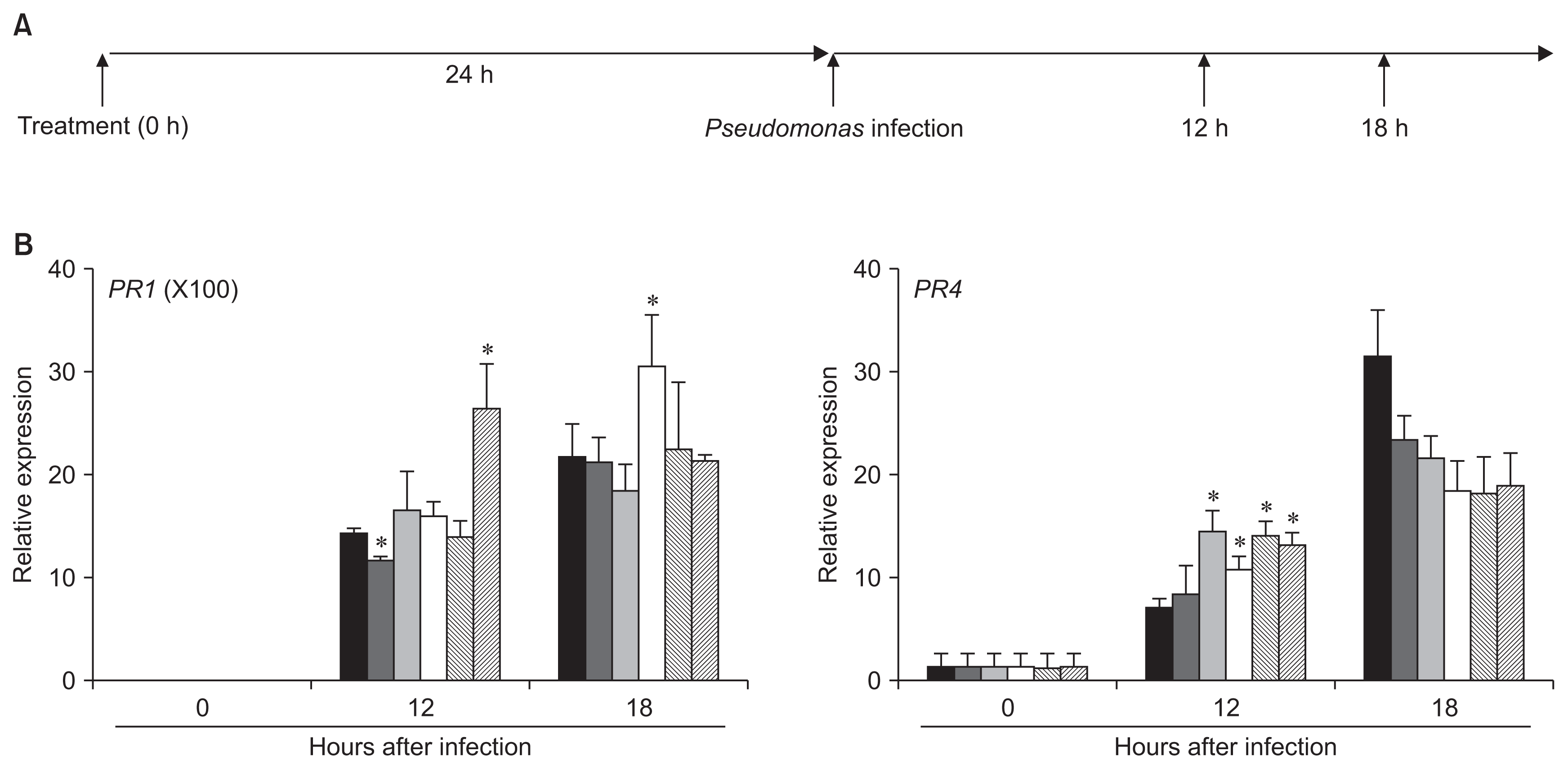
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






