



Banana (Musa acuminata L.), is a popular fruit and rich in nutrients, especially potassium (Kanazawa and Sakakibara, 2000). India is the largest producer of banana (30 million tonnes), holding 26.2% of the global share, with an export gross value of 3.48 billion USD (Ministry of Agriculture and Farmers’ Welfare, 2018). Nearly, 20-25% of fruits are lost due to poor postharvest management practices during various stages of the supply chain (Ministry of Agriculture, 2015). Such huge postharvest losses widen the gap between per-capita availability and supply which is estimated as nearly 50%. Ripening of banana is accompanied by a wide array of physiological and biochemical changes (Alkarkhi et al., 2011; Mohapatra et al., 2010). Being a climacteric fruit, it continues ethylene evolution, postharvest which makes the fruit deteriorate rapidly (Boudhrioua et al., 2003). Fungal pathogenic diseases in banana account for the major cause of postharvest spoilage at all stages of supply chain until consumption (Kasso and Bekele, 2018). Forty-two different genera of fungi are known to attack banana fruit during postharvest handling (Johnston and Jones, 1997). Anthracnose and stem-end rot caused by Colletotrichum musae and Lasiodiplodia theobromae, respectively are the most important fungal diseases affecting banana fruit (Abayasekara et al., 2013). These are opportunistic postharvest pathogens originating from quiescent infection and once established, the diseases are very difficult to manage.
The postharvest diseases are being managed through the application of fungicides as a dip or spray after harvest (Khan et al., 2001). Many fungicides are non-selective, leaving a considerable residue that affects the quality of fruit (López-Fernández et al., 2012). Hence, there is strong resistance from consumers who are alarmed by such residues, and its associated ill-effects on humans. Consequently, postharvest disease management has become quite challenging and there is increased need to find an alternate strategy to reduce the incidence of diseases using safe compounds that do not cause any health, safety or environmental issues.
Volatile compounds (organic and inorganic) are the products of secondary metabolism which can evaporate and disperse very rapidly into natural habitats and act as efficient chemical mediators. Many plant-based volatile organic compounds play a key role in the plant to plant communication (Baldwin et al., 2006), plant-insect and other defense interactions (Dobson, 2006; Heil and Silva Bueno, 2007). Similarly, bacterial volatile compounds have a broad impact on various organisms including plants, fungi, animals and bacteria (Sharifi and Ryu, 2018) acting as signaling compounds (Bailly et al., 2014), and plant defense elicitors (Ryu et al., 2004). Some of these volatile compounds act against postharvest pathogens and thereby the shelf life of fruit gets extended. There are several vapor treatments viz., 1-MCP, acetaldehyde, acetic acid, CO2 and hexanal that are used to delay the ripening of fruits (Baez-Sañudo et al., 2009; Fan et al., 2006; Gill et al., 2015; Myung et al., 2007; Song et al., 2007; Utto et al., 2008). The extension of shelf life of fruit is primarily attributed to the effective management of postharvest pathogens (Seethapathy et al., 2016; Sholberg, 2009).
Hexanal is a plant-derived biomolecule known to inhibit the phospholipase D (PLD) enzyme (lipoxygenase pathway) (Tiwari and Paliyath, 2011), and thus slow down the membrane deterioration (Paliyath and Droillard, 1992; Yuan et al., 2005) in fruits. Hexanal is categorized under generally recognized as safe and approved as a food additive by the U.S. food and drug administration, moreover, it is commercially available (Thavong et al., 2011). Hexanal formulations have been used as a preharvest spray (Anusuya et al., 2016; El-Kayal et al., 2017a; Kumar et al., 2018; Sharma et al., 2010), postharvest dip (El Kayal et al., 2017b; Jincy et al., 2017) and vapor (Sholberg and Randall, 2007) to extend shelf life of tropical and temperate fruits. Among the modes of delivery hexanal vapor treatment is a very simple, economically feasible and socially practicable method to extend the shelf life of fruits. Hexanal inhibits postharvest pathogens such as Botrytis cinerea and Penicillium expansum (Sholberg and Randall, 2005; Song et al., 2010; Thavong et al., 2011). In addition, it has been shown that the exposure of hexanal vapor enhances the volatile and aroma compounds of apple (Lanciotti et al., 2003; Song et al., 1996). Earlier, we have shown that hexanal formulation had no deterrence against honey bees and natural enemies in sprayed mango orchards (Mohan et al., 2017). Despite the fact that the effectiveness of hexanal vapor on spore germination and mycelial growth of fungal pathogens for postharvest diseases have been studied in temperate fruits, the work on tropical fruit like banana is less investigated.
The present study hypothesized that hexanal vapor inhibits the pathogenicity by triggering plant defense enzymes and thus provides resistance against postharvest pathogens. To address this hypothesis, the current work was undertaken with three specific objectives namely, (1) to assess the pathogenicity of Colletotrichum gloeosporioides and Lasiodiplodia theobromae after exposure to hexanal vapor, to gain insights into mechanisms involved in induced resistance (antioxidant enzyme activities and PLD) and to study the shelf life of banana fruit after exposing to hexanal.
Materials and Methods
Cultures and virulence
Two fungi, C. gloeosporioides and L. theobromae, were isolated from infected banana fruit exhibiting typical anthracnose and stem-end rot symptoms, respectively. Primary confirmations of pathogens were mainly based on morphological characters and further confirmed through PCR with internal transcribed spacer primers (Sangeetha et al., 2012). The pathogens were isolated by the tissue segment method on potato dextrose agar (PDA) medium (Rangaswami and Mahadevan, 1999). The infected banana fruit tissue portions were cut into small pieces of 1.0 cm2, surface sterilized with 0.1% mercuric chloride for 30 s, subsequently washed with sterile distilled water three times and dried over blotting paper. The surface-sterilized banana tissue segments were then placed on Petri plates containing 20 ml of PDA medium and incubated at room temperature (25 ± 2℃) for further fungal growth. The cultures were purified by a single hyphal tip technique and maintained on PDA slants at 4°C.
Hexanal vapor treatment
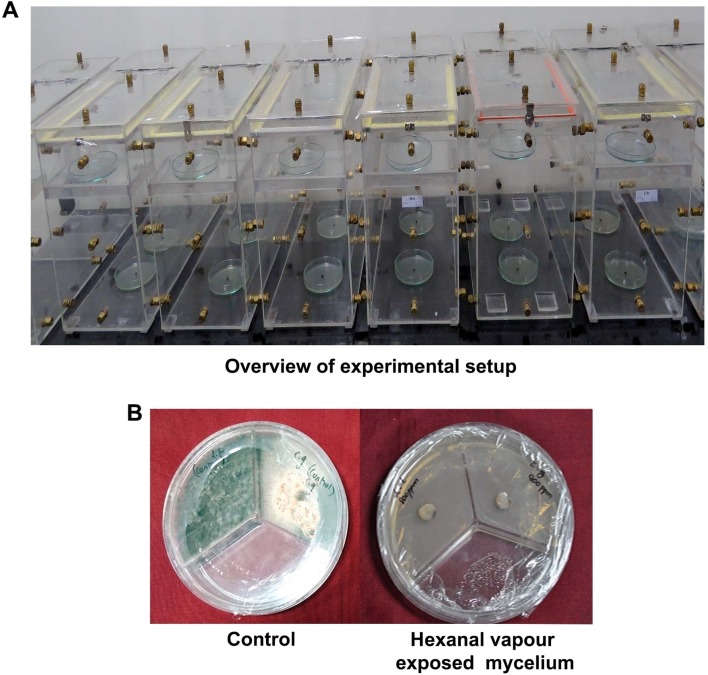
A plexiglass acrylic chamber measuring 45 cm length, 30 cm height, and 20 cm wide with a total volume of 27 l was designed to study the vapor treatment. The concentration of hexanal vapor was calculated by considering the chamber volume and liquid to gas conversion ratio based on Avogadro’s law (standard temperature and pressure, 273.15 K and 101.325 kPa, respectively). Thus to obtain various concentrations viz., 600, 800, 1,000, and 1,200 ppm of hexanal vapor, the volumes of hexanal liquid (98% purity, 0.8 g/ml density at 25°C; Sigma-Aldrich, St. Louis, MO, USA) required will be 101, 136, 170, and 204 µl, respectively. The required concentrations of hexanal liquid were taken up in Petri dish and placed inside the top phase of the chamber, which was laid out with a slot to stand.
Impact of hexanal vapor on fungal mycelial growth (in vitro)
The mycelial discs (5 mm diameter) of each fungal pathogen were taken from the periphery of an actively growing culture and placed at the center of a 90-mm Petri dish containing PDA medium and then placed on the bottom portion of the acrylic chambers. Various concentrations of hexanal as described earlier (to give 600-1,200 ppm) was pipetted out in Petri plates and placed inside the acrylic chamber (Fig. 1). The Petri plates containing fungal mycelium were exposed to each concentration at different time intervals (1, 2, 3, and 4 h) in different acrylic chambers. The control treatment had sterile water instead of hexanal vapor. The acrylic chamber was sealed with parafilm and placed at 25℃, with three replicates for each volume and each fungal species. After the completion of pre-defined exposure hours, the Petri plates were removed, sealed and incubated at 25℃ for further growth. After exposure to pre-defined time intervals the Petri plates were removed from the chamber, sealed and placed at 25°C. The colony diameter was measured daily until the control plates attained 90 mm diameter (full length). The inhibitory effect of each vapor treatment on mycelial growth was calculated as percent inhibition over the control.
Spore germination of C. gloeosporioides and L. theobromae (in vitro)
The effects of hexanal vapor on spore germination and germ tube elongation of C. gloeosporioides and L. theobromae were assayed according to Liu et al. (2009). Aliquots (50 μl) of both conidial suspensions at 2 × 106 spores/ml were plated on tripartite Petri dishes (90 mm diameter) with PDA medium containing different volumes of hexanal (0.36, 0.48, 0.60, and 0.72 μl) and all Petri dishes were incubated at 25°C. The respective controls were inoculated with sterile water instead of hexanal. Approximately, 10 spores of each pathogen were measured for germination rate within each replicate and the inhibition rate of spore germination was determined. Each treatment was replicated three times and the experiment was repeated twice.
Effect of hexanal vapor on banana diseases (in vivo)
Anthracnose and stem-end rot suppression efficacies of hexanal vapor treatment on banana fruit were also evaluated. The banana var. Grand Naine was randomly handharvested at 85% maturity from the university orchard, Tamil Nadu Agricultural University, Coimbatore (11.0°N latitude and 76.9E longitude), Tamil Nadu, India. Banana fruits with uniform size and maturity were collected and surface sterilized with sodium hypochlorite (1%) solution for 10 min. The pathogenic spores (5 × 105 cells) of L. theobromae and C. gloeosporioides were standardized using hemocytometer and 20 µl of the suspension was placed in the middle of the fruit after a small pinprick (Fig. 2). Then, the fruit was exposed to various concentrations (600-1,200 ppm) of hexanal solution placed in the top phase of the acrylic chamber and exposed to 1, 2, 3, and 4 h. Then the fruits were transferred to clean box and observed for disease symptoms at ambient temperature. The disease severity (Percent Disease Index [PDI]) was determined for both diseases on the 14th day of treatment. The experiment was replicated three times. For each experiment, we used one hand of banana fruits with six to seven fingers of the same maturity, weighing ~1.0 kg.
The treated and untreated fruit tissues were cut and fixed in 2.5% glutaraldehyde solution for 2-3 h. The fixed samples were then washed with an increasing concentration (step by step) of ethanol (0-100%) and dried. Post-fixation was done with 1% osmium tetraoxide according to Wang et al. (2008). The resulting samples were documented using scanning electron microscopy (SEM). The PDI was calculated based on Prabakar et al. (2005) with slight modification, where scale 0, no lesion development; scale 1, 1-25% fruit surface area covered with lesions; scale 2, 26-50% fruit surface area covered with lesions; scale 3, 51-75% fruit surface area covered with lesions; scale 4, > 75% fruit surface area covered with lesions.
Enzyme assays
Fruit samples were taken from the hexanal treated and control fruit with an interval of 1, 3, 6, 9, and 12 days after treatment. Exo- and endocarp tissue samples were collected, frozen in liquid nitrogen and stored at −80°C. Peroxidase (PO) activity (EC 1.11.1.7) was carried out as per the procedure described by Hammerschmidt et al. (1982) with slight modification in the reaction mixture total volume, where we used 2.5 ml instead of 2.9 ml.
For phenylalanine ammonia-lyase (PAL) activity, one gram of banana fruit samples was homogenized with 3 ml of ice-cold 0.1 M sodium borate buffer, pH 7.0, containing 1.4 mM of 2-mercaptoethanol and 50 mg of insoluble polyvinylpyrrolidone. The resulting extract was filtered and the filtrate was centrifuged at 20,000 ×g for 15 min at 4°C and the supernatant was used as the enzyme source. PAL activity (EC 4.3.1.5) was determined as the rate of conversion of L-phenylalanine to trans-cinnamic acid at 290 nm. Sample containing 0.4 ml of enzyme extract was incubated with 0.5 ml of 0.1 M borate buffer, pH 8.8 and 0.5 ml of 12 mM L-phenylalanine in the same buffer for 30 min at 30°C. The amount of trans-cinnamic acid synthesized was calculated using its extinction coefficient of 9,630/M/cm (Dickerson et al., 1984). Enzyme activity was expressed in fresh weight basis as nmol trans-cinnamic acid/min/mg of plant tissue.
For glucanase (EC 3.2.1.39) activity, 1g tissue of banana peel and pulp were homogenized in 5 ml of 0.05 M sodium acetate buffer (pH 5.0) and the homogenate was centrifuged at 20,000 ×g for 10 min at 4°C and the supernatant was used as enzyme source. β-1,3-glucanase activity was assayed by the laminarin-dinitrosalicylic acid method (Pan et al., 1991). The reaction mixture consisted of 62.5 µl of 4% laminarin (Sigma-Aldrich) and 62.5 µl of enzyme extract. The reaction was carried out at 40°C for 10 min. The reaction was stopped by adding 375 μl of dinitrosalicylic acid and heated for 5 min in boiling water, vortexed and its absorbance was measured spectrophotometrically at 500 nm (SPECORD 210 PLUS, Konrad-Zuse-Str-Jena, Germany). The enzyme activity was expressed as µg glucose released/min/mg of the sample.
Phenolic compounds were assayed as per the method of Liu et al. (2008) with some modifications. The samples (0.5 g) were homogenized with 5 ml ice-cold 1% HCl-methanol solution and then centrifuged at 12,000 ×g for 10 min at 4°C. The supernatant was removed, absorbance measured at 280 and 325 nm in a spectrophotometer (SPECORD 210 PLUS). Gallic acid was used as a standard.
Determination of PLD activity
The fresh tissues of banana were ground using 0.2 M phosphate buffer (pH 7.0) and the PLD enzyme activity was quantified using PLD assay kit (EC 3.1.4.4., Sigma-Aldrich) following the procedure described in Jincy et al. (2017).
Evaluation of hexanal vapor on the pathogenesis of fruit peel and pulp
To determine the effects of hexanal treatment on fruit peel and pulp morphology, as well as the effect of pathogen inoculation, samples were collected on the 6th day after treatment (color break stage in control fruits). The peel and pulp samples were collected from the middle portion of the fruit and immediately dipped -20℃ ethanol for 3 s and stored at −80℃ until further analysis. At the time of analysis, the sample was affixed to a carbon stub and vacuum desiccated for 2 min to remove excess moisture. The peel and pulp samples were examined using a scanning electron microscope (Quanta 250, FEI, Hillsboro, OR, USA) with an Everhart-Thornley Detector detector. The SEM was operated in a vacuum, 10 kV, with a spot size of 3.5 and a pressure of 60 Pa. The images were recorded at 500× and 1,000× magnifications.
Shelf-life studies
Fruit samples were drawn once in three days from control and treated, till the fruit get completely spoilt to assess the shelf life using physiological, biochemical and quality parameters as repeated earlier (Anusuya et al., 2016).
Statistical analysis
The data were statistically analyzed using the IRRISTAT version 92 (International Rice Research Institute Biometrics unit, Manila, Philippines) developed by the International Rice Research Institute Biometrics unit, the Philippines. Prior to statistical analysis of variance (ANOVA), the percentage values of the disease index were arcsine transformed. Data were subjected to ANOVA at two significant levels (P < 0.05 and P < 0.01) and means were compared by Duncan’s multiple range test (Supplementary Tables 1-8).
Results
Hexanal vapor treatment on mycelial growth and spore viability
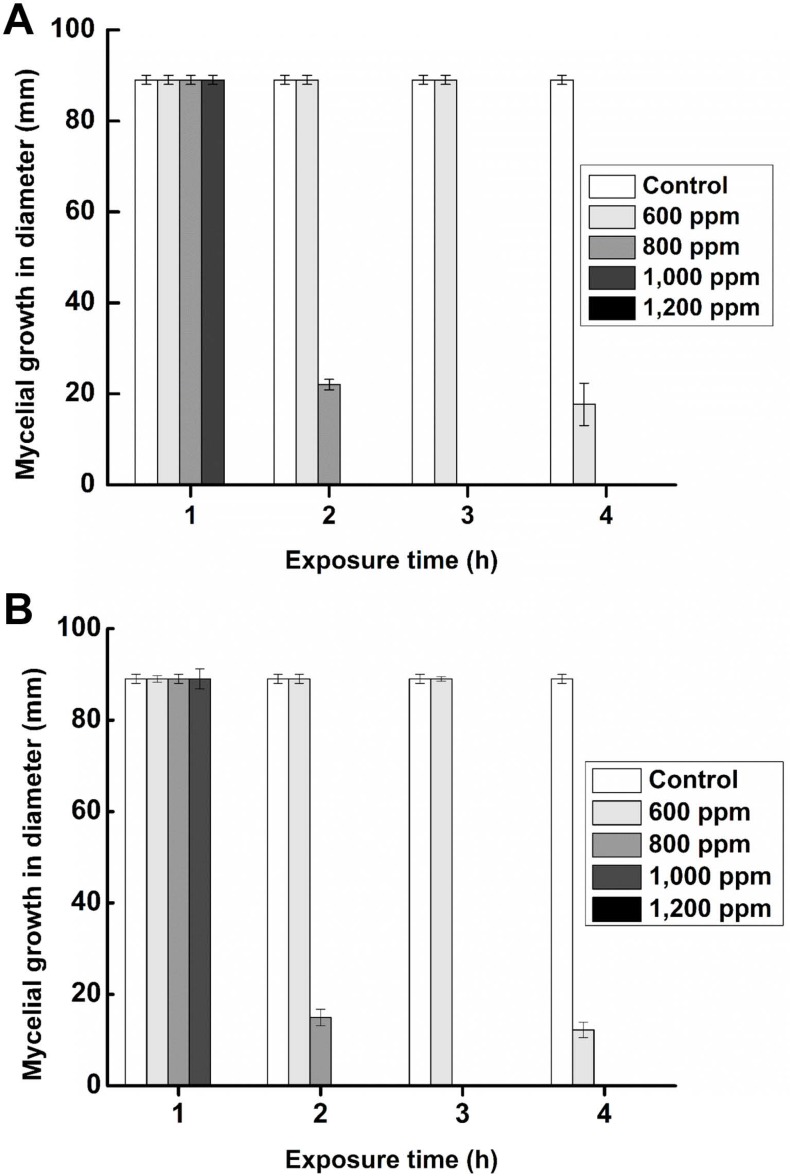
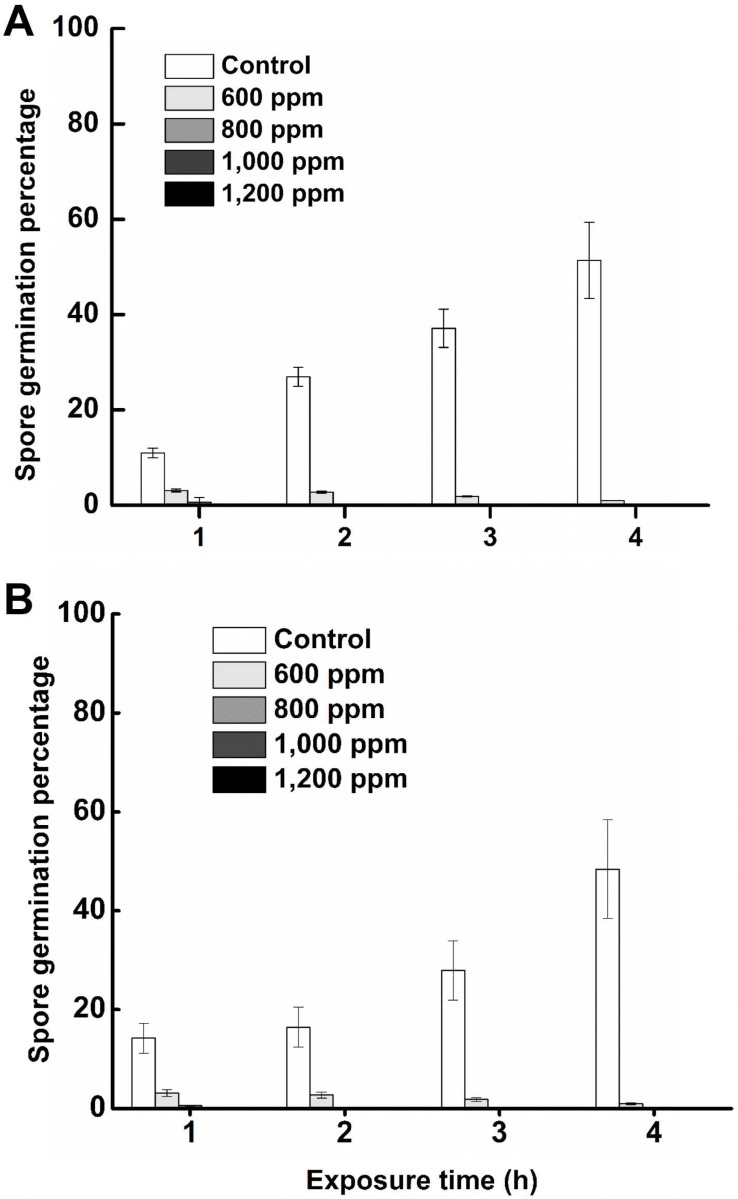
The pathogens exposed to varying concentrations of hexanal vapor (600 to 1,200 ppm) under in vitro conditions and the maximum mycelial growth in control plates were attained after 7 days of inoculation. Exposure to hexanal at 1,000 or 1,200 ppm for one hour resulted in the complete inhibition of mycelial growth in both. At a lower concentration of hexanal, such as 800 ppm it required longer exposure time (3 h) to achieve complete inhibition (Fig. 3). Spore viability of C. gloeosporioides and L. theobromae decreased with increasing concentrations and duration of exposure to hexanal. At 800 ppm (and higher) of hexanal vapor, spore viability decreased rapidly and germination was totally inhibited after an hour of exposure (Fig. 4). Hence 800 ppm was used in commercial fruit packaging studies/assay due to complete inhibitory effect at low concentrations as well as the economic benefit gained through minimal usage of a volatile compound.
Effect of hexanal vapor on lesion development and shelf life
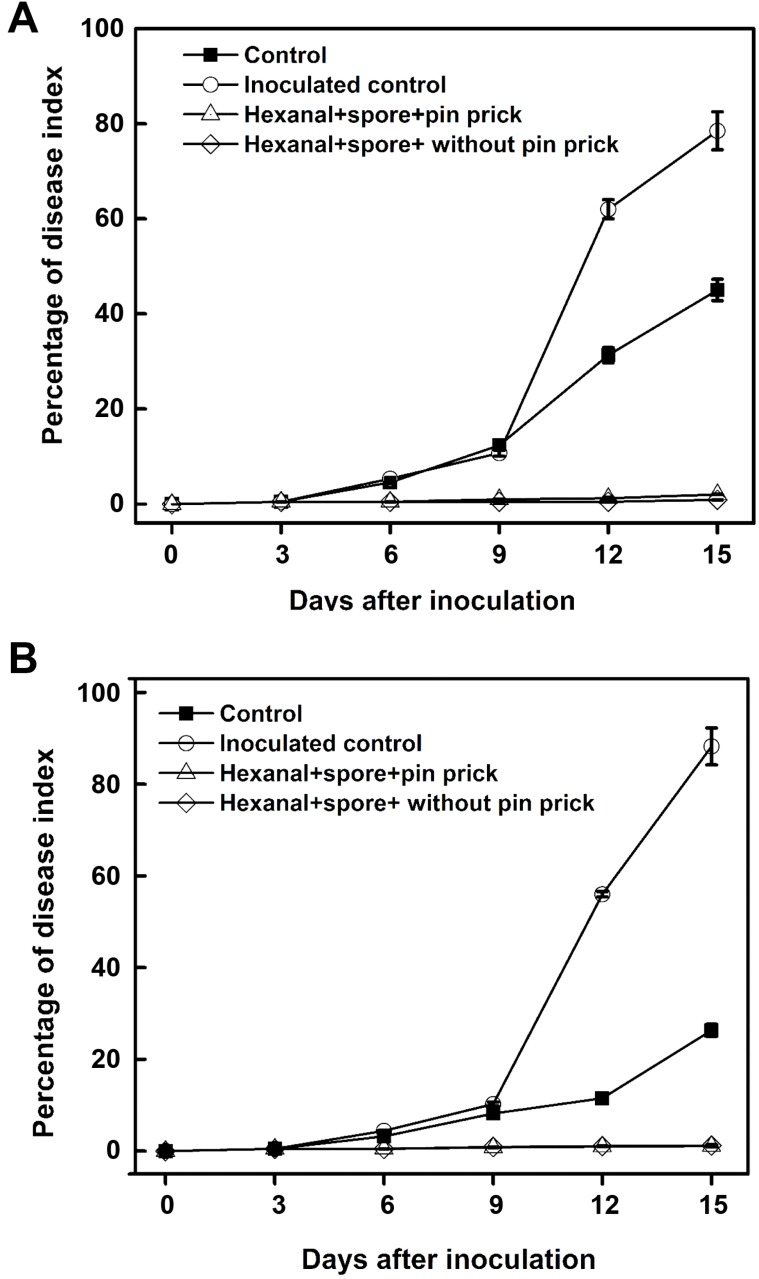
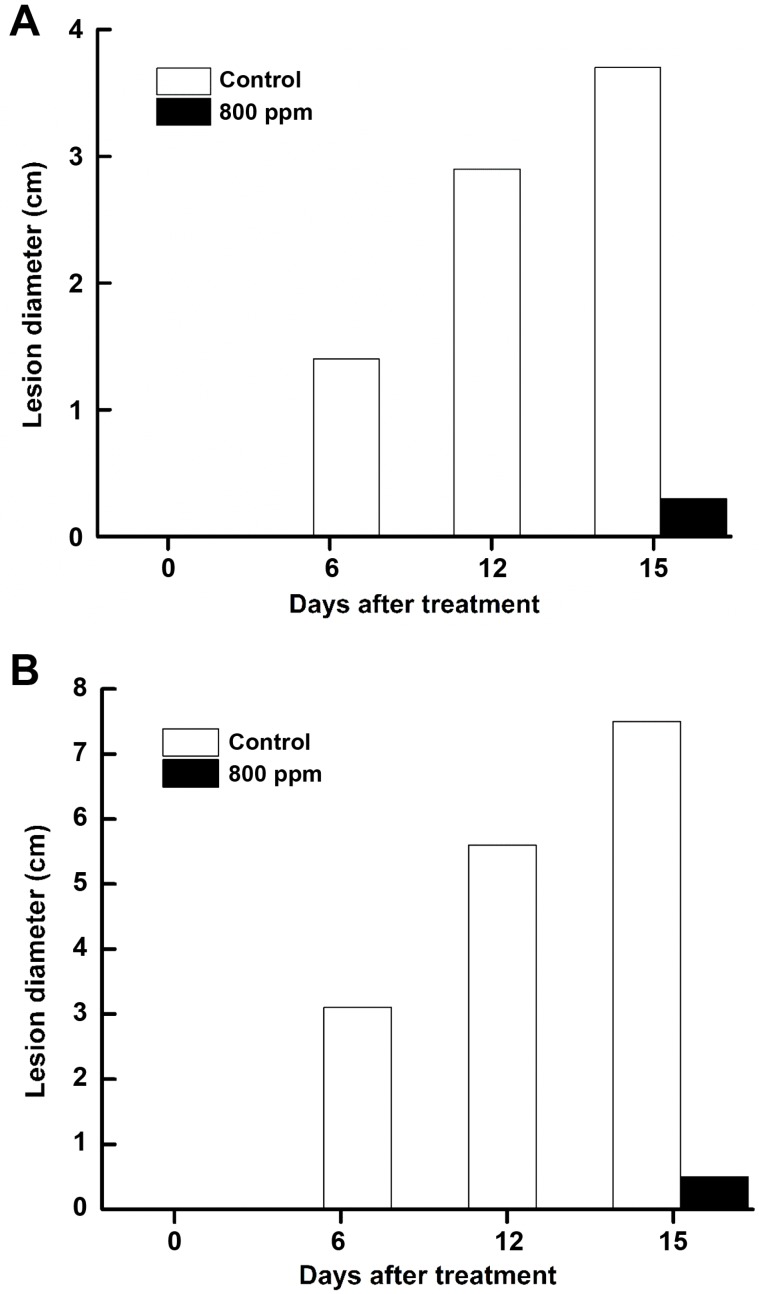
In general fruits stayed without any noticeable lesion for 6 days and thereafter lesions were more prominent. Fruits that were exposed to 800 ppm for 3 h had registered the lowest incidence of stem-end rot (1.61%) and anthracnose (1.12%) on 15th day of observation. On the other hand, infection started becoming visible on 6th day in control fruits and progressed steadily thereafter. On the 15th day incidence of stem-end rot and anthracnose reached 78.5% and 88.2% respectively and the fruits were lost for any practical purposes (Figs. 5 and 6). Correspondingly, the treated fruits stayed well until 14 days, fit for marketing/consumption. Hexanal vapor exposed fruit had the smallest lesion size of 0.3 and 0.5 cm for stem-end rot and anthracnose, respectively at 15th day. The corresponding lesion sizes measured in control fruit were 3.7 and 7.5 cm (Fig. 7). We observed the disease incidence in healthy control even after surface sterilization (Fig. 5), appears to be a normal case in banana fruits. This might be due to the endemic as a latent form of pathogen propagules which is carried from green fruit and expressed symptoms during ripening. Our scoring (PDI formulae) was based only on true disease symptoms rather than normal discoloration.
Changes in the PO, PAL, and glucanase activities in banana fruit
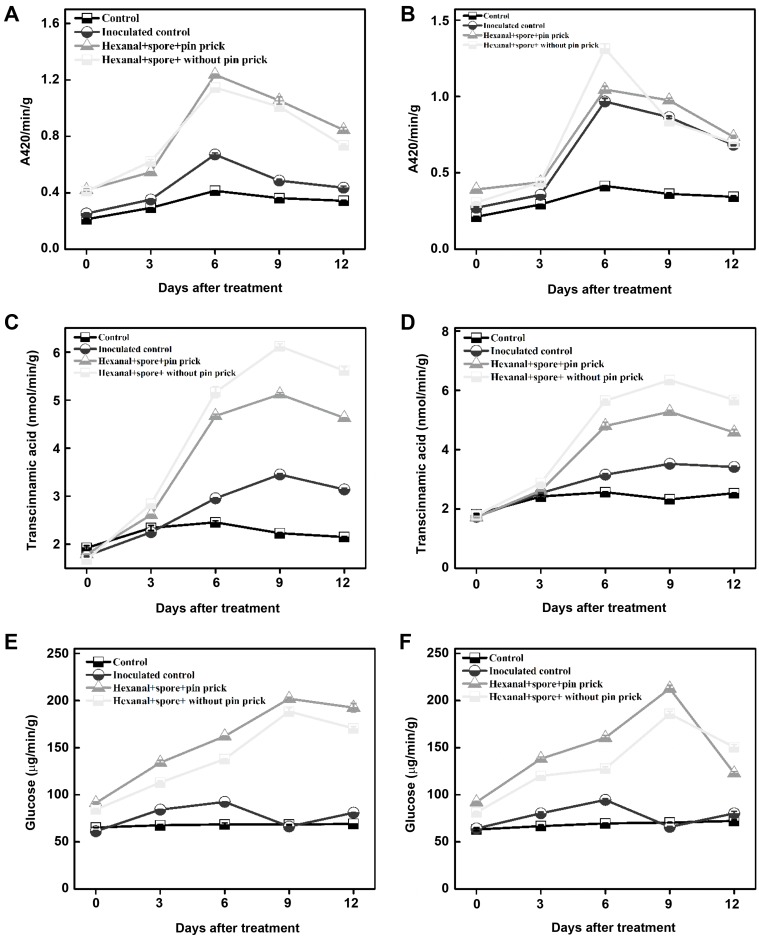
The PO activity had transiently increased till seven days and declined thereafter regardless of treatment. Hexanal treatment (800 ppm) and pre-inoculation with both pathogens (C. gloeosporioides, L. theobromae) had significantly higher PO activities (1.314 and 1.312 changes in absorbance/min/g of fresh tissue, respectively) than control (0.414 changes in absorbance/min/g of fresh tissue) at the 6th day of treatment and decreased gradually thereafter (Fig. 8A and B, Supplementary Tables 3 and 7). Similarly, PAL activity also increased due to hexanal till 9th day (6.12; 6.342 nmol of trans-cinnamic acid/min/g of fresh tissue respectively), compared to control (2.23 nmol of trans-cinnamic acid/min/g of fresh tissue). PAL declined thereafter regardless of treatments (Fig. 8C and D). The β-1,3-glucanase also increased when exposed to hexanal vapor at 800 ppm till 9 days (212 and 201 µg of glucose/min/g of fresh tissue) in comparison to control (68 µg of glucose/min/g of fresh tissue) and then decreased thereafter (Fig. 8E and F). Together these data indicate that hexanal induces significant biochemical changes in banana that are associated with shelf life and defense against postharvest fungal pathogens.
Changes in the phenolic content of banana fruit
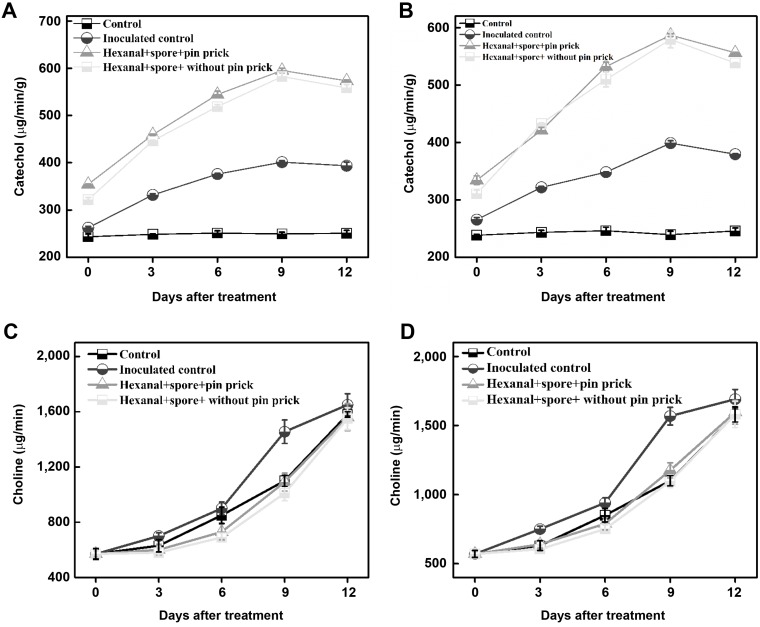
Accumulation of phenolics was observed in pre-inoculated banana fruit treated with hexanal vapor (800 ppm) and registered the highest value on the 9th day after treatment. The hexanal vapor treated banana fruit pre-inoculated with L. theobromae and C. gloeosporioides recorded higher phenolics (595.2 and 587.1 catechol equivalents/g of fresh tissue) than control (249.3 and 239.3 catechol equivalents/g of fresh tissue). The control fruit had increased phenolics up to 6 days after treatment and declined thereafter (Fig. 9A and B).
Changes in the PLD activity
The activity of PLD in hexanal vapor treated fruits was lower than control during the entire period of observations (0 to 12 days). The PLD activities were lower in hexanal vapor treated fruit regardless of inoculation with either pathogen (Fig. 9C and D).
High-resolution imaging
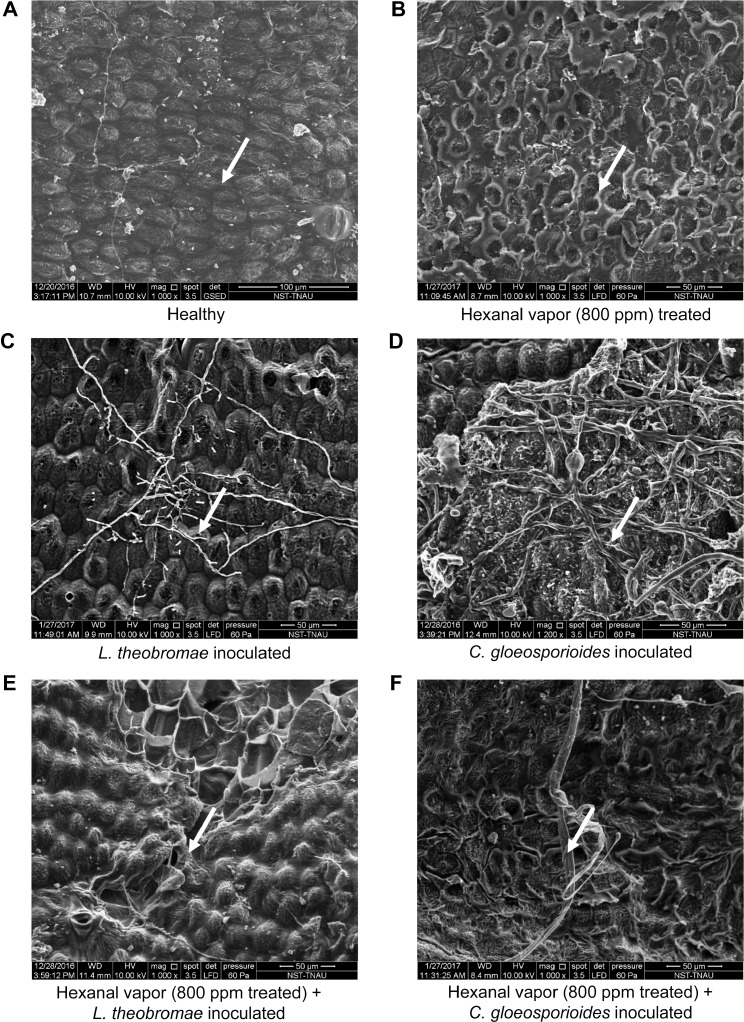
The high-resolution images of SEM exhibited colonization and penetration of L. theobromae and C. gloeosporioides on the surface of the control fruit. On the other hand, hexanal vapor exposed fruit had shown to have lignifications on the surface of the fruit and no colonization was observed on the 9th day of treatment (Fig. 10A-F).
Discussion
Infection by postharvest pathogens may occur before harvest, during the growing season or after harvest, or during the handling and transport. Once the pathogens invaded the tissues, it is hardly possible to prevent the spread of the disease as the fruits start to ripen and will be consumed within a short time after. In many cases, the postharvest diseases are rarely recognized with obvious symptoms during the growing season, as the pathogen remain latent during this phase. Hence controlling latent infections in the postharvest environment of fruits is very important (Caruso and Ramsdell, 1995). Since anthracnose and stem-end rot cause extensive damage during postharvest, any preventive treatment has to be integrated well before the pathogens start infecting the fruit tissues.
At present fungicides are used at postharvest to control such infections (Stoytcheva, 2011) and there is an immediate need to find alternative solutions. Some of the natural volatile compounds such as cinnamaldehyde and benzaldehyde significantly inhibited the mycelial growth and reduced disease incidence of some postharvest diseases such as stem-end rot, anthracnose, gray mold and brown rot of fruits during postharvest storage (Sivakumar et al., 2001; Wilson et al., 1987). Hexanal can be used as another volatile to control the proliferation of postharvest pathogens. The exposure to hexanal vapor causes the hyphal cell wall membrane disruption (Fallik et al., 1998) and distortion of cell wall components, plasma membrane in conidia (Arroyo et al., 2007). In addition, it has been shown that the hexanal vapor reduced 94% of spore viability in Penicillium expansum (Fan et al., 2006).
In this study, we have shown sufficient evidence that hexanal can be an effective treatment to control the postharvest pathogens and to extend the shelf life of banana fruits during storage. Treating banana fruits with 800 ppm of hexanal had increased the postharvest shelf life to 15 days (Figs. 5 and 6). Such an extension is most likely due to the inhibition of spore germination and mycelial growth of these fungi by hexanal exposure for just 3 h (Figs. 3 and 4). Hexanal has been shown to have similar inhibitory effect on other postharvest pathogens such as B. cinerea and M. fructicola (Song et al., 2007). Recently, Seethapathy et al. (2016) have shown that the hexanal treated mango fruit volatile profiles were uniquely different in terms of additional compounds detected that are known to have fungistatic effects. Perhaps hexanal may have such an indirect effect by stimulating the release of such fungistatic volatiles from banana as well. However, this needs to be explored further to draw any conclusions
Appearance of disease incidence in later stages of ripening in the surface-sterilized control is likely due to latent pathogens that become virulent during ripening. It should be noted that the scoring was based only on true disease symptoms rather than normal discoloration which generally appears in the banana. Earlier studies have shown that hexanal vapor treatment in peach fruits inhibited the fungal growth such as C. gloeosporioides (Song et al., 2007), M. fructicola, and M. laxa in peaches (Baggio et al., 2014) and mold such as Penicillium expansum in pear fruits (Neri et al., 2006). Apples exposed to 900 μl/l hexanal vapor for 48 h showed signs of phytotoxicity, expressed as surface browning (Fan et al., 2006). This could be a much higher dose and longer duration as our studies indicate it to be effective 5.02 µl/l for 3 h without any phytotoxicity symptoms. These results suggest that hexanal is quite effective at low dose that can be adopted for commercial level postharvest treatment.
In plants, the defense enzymes such as PO, PAL, glucanase, and chitinase are induced as a first line of defense against pathogen infection (Anand et al., 2007; Barber and Mitchell, 1997; Dixon et al., 1995). PO, a key enzyme involved in various defense-related processes includes cross-linking of phenolics-glycoproteins, polysaccharidesmonomers, and hypersensitive response (Thakker et al., 2013). In addition, the deposition of phenolic materials on the cell membranes acts as a barrier for the development of the pathogen (Agrios, 2005; Thakker et al., 2013). Phenolic compounds enhance the mechanical strength of plant cell walls and also act as defense molecules against pathogen infection (Ganeshamoorthi et al., 2008; M’Piga et al., 1997). Increased PAL levels were observed during pathogen infection, other biotic and abiotic stresses (Lafuente et al., 2003). β-1,3-glucanase is primarily involved in the cell wall modifications tailored towards microbial defense (Beffa et al., 1996; Smart, 1991). In general, β-1,3-glucanase enzymes degrade both plant and pathogen cell wall and derivatives arose from cell wall degradation act as elicitors in defense response (Bowles, 1990). Hexanal induced elevation in β-1,3-glucanase, PAL, PO, and phenolic activity in treated banana fruits could be the reason for the increased resistance against pathogens.
Fruit softening during the ripening process is associated with loss of membrane integrity attributed by families of enzymes including cellulase, glucanase, polygalacturonase, and phospholipases (Payasi et al., 2009). PLD is a key enzyme involved in the hydrolysis of phospholipids, which are the primary backbones of the cellular membrane and increased PLD activity is observed during fruit ripening in various crops (Paliyath and Droillard, 1992; Yuan et al., 2005). Hexanal inhibits the PLD enzyme thereby increasing the shelf life of fruits. Our result shows that the PLD levels increased during ripening as well as due to fungal inoculation in the control fruits, while, hexanal lowers the PLD up to the 9th day of storage (Fig. 9). This results in the extension of shelf life. This clearly shows that the membrane integrity was maintained until 9th day by inhibiting PLD in the treated fruits which likely inhibited fungal growth due to hexanal treatment. Recently it was shown that hexanal based nanoformulation treated banana fruits exhibited extra firmness, shininess, increased total soluble solids, sugar levels indicating improved quality during storage (Venkatachalam et al., 2018) as observed in the current study.
The primary mechanism of action of hexanal on C. gloeosporioides and L. theobromae is not fully understood. Based on our evidence it is possible that keeping the membrane intact as a result of hexanal treatment may have contributed to a type of physical resistance (confirmed by SEM), coupled with elevated levels of defense molecules described previously, contributing to biochemical resistance. Thus the cumulative effects render the hexanal treated fruits with an extended shelf life and thus contributing to lessen postharvest waste.
The high level of disease control achieved when naturally infected banana fruit treated with hexanal, suggests that it can penetrate tissues to a certain extent. It has been shown that the whole apples inoculated with P. expansum at 22℃ and 4℃ exposed to hexanal vapor reduced the decay (Fan et al., 2006), indicating that it could be applied commercially at low temperature. Although we did not test temperature effects in the present study, it is possible to increase hexanal vapor concentration at lower temperatures by using heat-vaporization. The volatile nature of hexanal and its potential ability to penetrate stacked commodities would be an added advantage to expose the fruits during transport and storage. The potential use of hexanal as a tool for postharvest shelf-life extension is promising from our current studies, although large scale studies might be needed to figure out the economics more accurately. The fact that hexanal vapor is as effective as a liquid opens up its use for extending shelf life in other soft fruits where using a liquid form of protection is not preferred. Further using such external products as a vapor also limits its exposure to the fruits as well as environment thus making it ecofriendlier.
Hexanal is a natural volatile and had significant inhibitory effects on postharvest pathogens and it can be easily incorporated into supply chain management (Supplementary Tables 1-8). Treatment with hexanal vapor at 5.02 µl/l for 3 h substantially reduced the infection and decay of banana fruit. The use of hexanal as an antimicrobial fumigant is a promising alternative for chemical treatments.
Electronic Supplementary Material
Supplementary materials are available at The Plant Pathology Journal website (http://www.ppjonline.org/).


 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement
Supplement Print
Print






