

Introduction
Pasteuria penetrans has potential for its development as a biological control agent of economically important root knot nematodes (Sayre and Starr, 1985). These hyperparasites start their infection by adhering to the cuticle of infective juveniles (J2s), then penetrate the body wall to enter the host root, usually after the nematode has set up a feeding site (Chen et al., 1997). They either act as restraining the nematode migration toward the roots or by reducing their reproduction (Brown and Smart, 1985; Davies et al., 1988).
Adhesion of spores to the cuticle plays a vital role in the activity of the pathogen when it is used as a biological nematicide (Stirling et al., 1986). Attachment studies of isolates of Pasteuria to different populations and strains of RKN showed a high degree of variation and it is host specific (Davies et al., 1988; Espanol et al., 1997).
This obligate parasitic bacterium exhibits host specificity at two stages in the nematode life cycle; firstly, endospore isolates differ in their ability to encumber the J2s of particular species and strains of root-knot nematodes (Davies, 2005; Davies et al., 2001) and second, endospore populations differ in adhesion to different life stages of the same strain as exemplified by differential attachment to males (Davies and Williamson, 2006).
Identification of RKN is a prerequisite to develop an effective cropping system, resistant cultivars and biocontrol agents (Cenis, 1993). Molecular approaches have made significant contribution to the detection and identification of RKNs (Randig et al., 2002; Ward et al., 2004). Polymerase chain reaction (PCR) based techniques are a strong diagnostic tool for accurate detection of pathogens and have also been widely used for the identification of nematodes (Han et al., 2004; Henson and French, 1993; Martin et al., 2000; Schaad and Frederick, 2002). However, no comprehensive study has been conducted for genetic variability assessment in RKN in Pakistan using molecular tools. Hence, the present study was aimed to in-vitro encumbrance of P. penetrans with different RKN populations at different temperatures and concentrations, and to identify intra-Meloidogyne genetic variability in RKN populations which have better encumbrance with P. penetrans, collected from different ecological regions of Pakistan through PCR-RFLP of an intergenic region between cytochrome oxidase subunit II (COII) and 16S ribosomal mitochondrial genes.
Materials and Methods
Survey for the occurrence of different root knot nematodes in vegetable production areas of Punjab-Pakistan
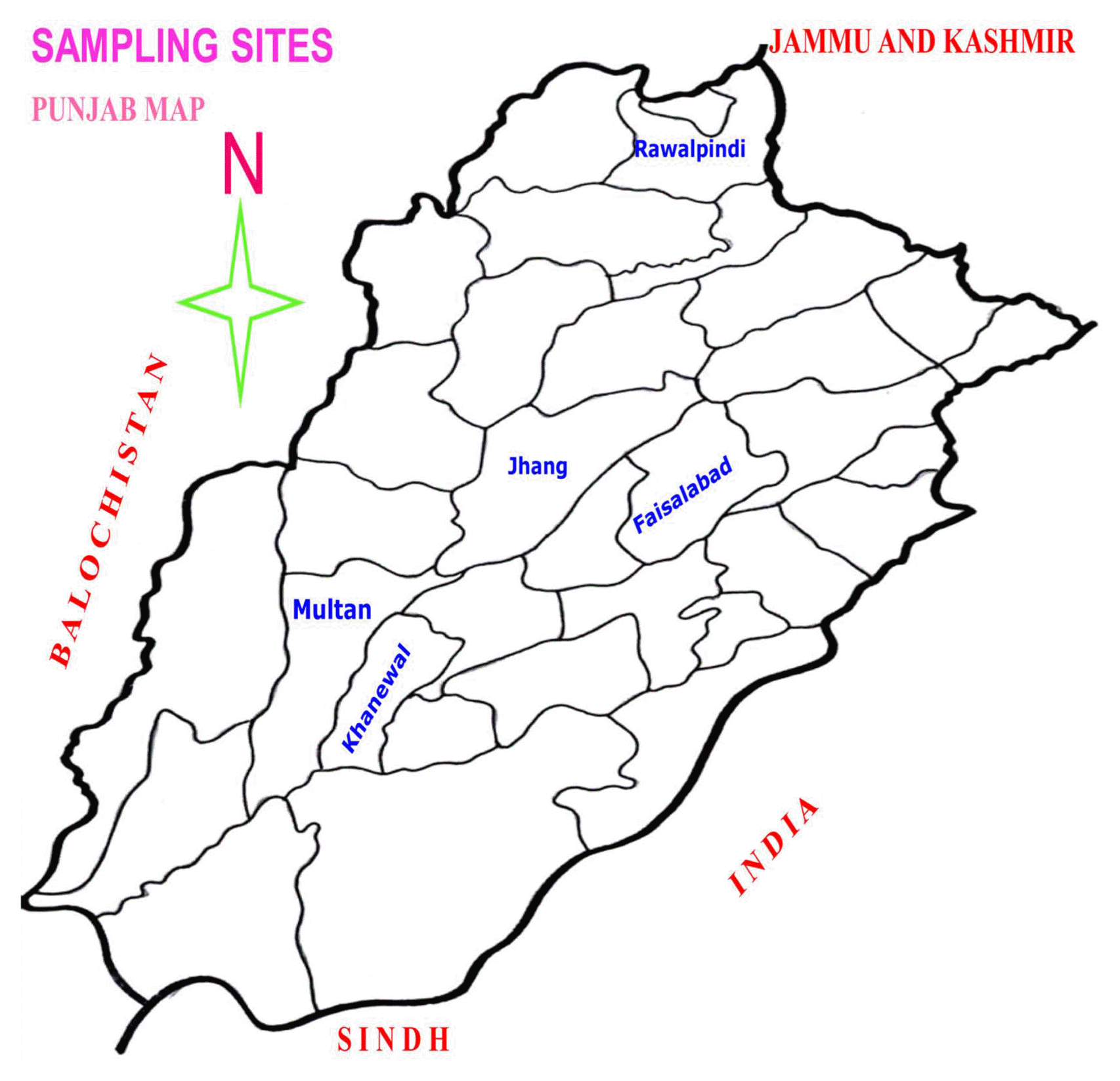
A systematic survey was conducted for a reliable estimation of infestation of root knot nematode major vegetable production area of five Districts of Punjab Province [Faisalabad, Jhang, Khanewal, Multan and Rawalpindi (Fig. 1)]. From each sampling site ten samples were collected at random. Root samples were carefully lifted with trowel up to 15-20 cm depth from the rhizosphere of tomato and cucumber plants together with approximately 1 kg of adhering soil. Samples were put in polythene bags and data on host, locality and soil type etc. was recorded.
A total of 700 samples were collected from two vegetable hosts i.e. tomato and cucumber (350 samples from each host). These samples were transported in Nematology Laboratory of Plant Pathology Department, University of Agriculture, Faisalabad-Pakistan and were stored in refrigerator at 5°C (40°F) until processing.
In vitro encumbrance of P. penetrans with different root knot nematode populations at different temperatures and concentrations
KN populations used for encumbrance test
A total of 116 populations of RKN (two population/sampling site) were selected for their encumbrance to two isolates of Pasteuria (PP-3 and PP-J). The selection of these populations was made on the basis of RKN incidence in different sampling sites of five District of Punjab, Pakistan on two hosts (tomato and cucumber). During survey 27 out of 35 sampling sites were infested with RKN in case of tomato while in case of cucumber 31 out of 35 sampling sites were infested with RKN, hence 54 RKN populations (27 × 2 = 54, two RKN populations/sampling site) were selected from tomato and 62 RKN populations (31 × 2 = 62, two RKN populations/sampling site) were selected from cucumber.
Nematode inoculums
All RKN populations were purified by single egg mass culture and maintained on susceptible tomato cv. Money maker throughout the experiment. Root system bearing egg masses of each Meloidogyne population was washed, cut into pieces and treated with 1% Sodium hypochlorite (to facilitate the release of eggs from egg masses) two days before setting experiment (Hussey and Barker, 1973). The resulting egg suspension was placed in extraction dishes and incubated at 28°C for hatching. The newly hatched juveniles were collected after 2 days of incubation period and used in attachment studies.
Pasteuria endospore concentrations
Dry root powder (100 mg) containing mature endospores of two isolates of Pasteuria (PP-3 and PP-J) were grinded with a pestle and mortar in two ml of water. More water was added and sieved through 38 μm sieve (Stirling and Wachtel, 1980) and two suspensions of Pasteuria endospores were prepared in water (104 and 105 spores/ml) using Haemocytometer. Dilutions of the suspension were made using the following formula;
One ml of the spore suspension was pipetted to each Petri dish (5 cm in diameter) containing one ml of nematode suspension (50 J2s/ml). To facilitate the counting suspension was diluted by adding one ml of water. Each treatment was replicated ten times, completely randomized and incubated at 25 ± 2°C. Similarly, another set was prepared and incubated at 30 ± 2°C in an incubator.
Estimation of spore attachment
The Petri dishes were checked after one hour of exposure time and the number of spores attached/J2s on first 10 nematodes was counted under the microscope at 400× magnification. The mean of ten counts was calculated as one observation for each replication. Same procedure was repeated for each nematode population isolated from different hosts.
Assessment of genetic variability among different populations of root knot nematodes using PCR-RFLP of CO II and large subunit of rRNA gene (lrRNA) of the mitochondrial genome
Nematode populations used in PCR
A total of 19 RKN populations (from tomato and cucumber) were subjected to PCR based analysis of intra-Meloidogyne genetic variability. The populations were selected on the basis of their encumbrance to two isolates of P. penetrans in the in-vitro encumbrance experiments (Table 1). Seventeen RKN populations were selected on the basis of their maximum encumbrance. Two populations which produced minimum encumbrance as compared to all the Meloidogyne populations were also included in the study.
DNA extraction
Total genomic DNA was extracted from single J2 using worm lysis buffer (WLB) [50 mM KCl, 10 mM Tris pH 8.3, 2.5 mM MgCl2, 0.45% Tween 20 (Sigma-Aldrich, UK), 0.01% gelatine, 60 μg/ml proteinase K (Fermentas, USA)]. An individual J2 was placed in 10 μl of WLB on a sterilized glass microscopic slide, cut into half with sterile micropipette tip, pressed, and lysate was transferred into 0.5 ml PCR tube. The tubes were frozen at −20°C for 10 min, incubated at 65°C for one hour followed by 95°C for 10 min and then immediately placed on ice. An aliquot of 5 μl of the suspension was used in PCR.
Polymerase chain reaction (PCR)
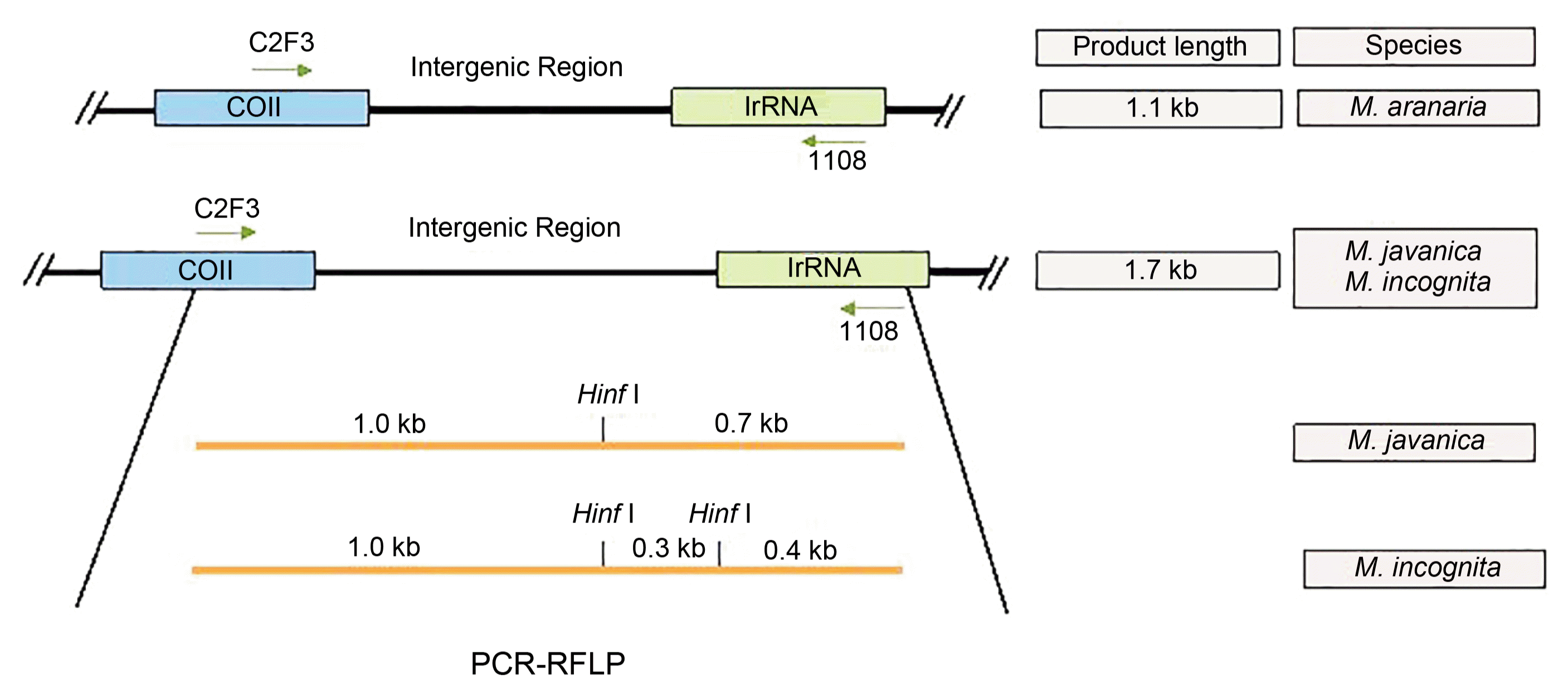
The molecular characterization was conducted following protocol reported by Powers and Harris (1993). A fragment of intergenic region between COII and large subunit of rRNA gene (lr-RNA) of the mitochondrial genome was amplified using forward primer C2F3 (5′-GGTCAATGTTCAGAAATTTGTGG-3′) and reverse primer 1108 (5′-TACCTTTGACCAATCACGCT-3′). Detailed description of primer design is given in Fig. 2. Polymerase chain reaction was performed in 20 μl reaction volume in 0.2 ml PCR tubes in a thermal cycler (Mastercycler gradient, Eppendorf Germany). Composition of the reaction mixture and temperature cycles is given in Table 2 and 3, respectively.
Gel Electrophoresis and Analysis
The PCR products were resolved on 1.5% agarose gel. In brief, agarose powder was dissolved completely in by sprinkling slowly over the stirring 1× Tris-Borate-EDTA (TBE) buffer and melted completely by heating in microwave oven in short intervals avoiding excessive foam formation and spitting out of the flask. Ethidium bromide was added at 0.5 μg/ml after cooling the solution to ~60°C and the solution was poured into the gel casting tray. After polymerization, the gel was placed in electrophoresis tank. The 6× DNA loading dye was added to the products and 10 μl of the mixture was pipetted into wells. First and last well of the gels were loaded with 1 kb DNA ladder (Fermentas, USA) for determination of exact size of amplicons. Electrophoresis was performed in 1× TBE buffer at 80 V for 1 h. Gels were visualized and documented using gel documentation system (NYXTECH-NIK, UK).
Restriction analysis of PCR products
The PCR products of 1.7 kb in size were further subjected to restriction analysis to discriminate between M. incognita and M. javanica species through digestion with Hinf I enzyme. A 20 μl restriction reaction was assembled using 10 μl of amplified product, 7 μl of nucleases free water, 2 μl of 10× buffer R [10 mM Tris-HCl (pH 8.5), 10 mM MgCl2, 100 mM KCl, 0.1 mg/ml BSA] and 1 μl (10 units) of Hinf I enzyme (Fermentas, USA). Contents were gently mixed, briefly centrifuged and incubated at 60°C for two hours. An aliquot of 10 μl of the restriction mixture was resolved on 1.5% agarose gel, stained with ethidium bromide and visualized through gel documentation system as described above.
Results
In vitro encumbrance of P. penetrans with different RKN populations at different temperatures and concentrations
Encumbrance of PP-3 with different RKN populations at different temperatures and concentrations isolated from tomato
A great variable response was observed when PP-3 encumbered with different populations of RKN at two different temperatures (ambient and 30 ± 2°C) and concentrations (104 and 105 spores/ml). The encumbrance level of 54 populations of RKN isolated from tomato varied significantly (Table 4). Interspecific and intraspecfic variability in encumbrance of PP-3 with different populations of RKN was demonstrated. The encumbrance of PP-3 at concentration of 104 spores per ml at ambient temperature was as low as below ten spores attached to different populations of RKN collected from five District of Punjab. Rate of spore attachment at 105 spores per ml was approximately double that at 104 spores/ml (Table 4). At 30 ± 2°C, the encumbrance level also increased in all the RKN populations. The results revealed that with the increase in temperature and spore concentration the mean encumbrance level of Pasteuria with J2 cuticle increased. At least one population of RKN was selected from each District having maximum encumbrance with PP-3. These populations were FSD/T8 (Faisalabad), JNG/T11 (Jhang), KHW/T6 (Khanewal), MUL/T3 (Multan) and RWP/T3 (Rawalpindi).
Encumbrance of PP-J with different RKN populations at different temperatures and concentrations isolated from tomato
The same 54 populations of RKN responded differently when encumbered with PP-J isolate of Pasteuria. Its attachment with different populations of RKN at two different temperatures and concentration was almost double than PP-3. The RKN populations which gave maximum encumbrance with PP-J were different from those RKN populations which showed maximum encumbrance with PP-3 (Table 4). Only FSD/T8 (Faisalabad) population showed maximum encumbrance with PP-3 as well as PP-J. The other four populations which gave maximum encumbrance were JNG/T3 (Jhang), KHW/T2 (Khanewal), MUL/T10 (Multan) and RWP/T7 (Rawalpindi).
Encumbrance of PP-3 with different RKN populations at different temperatures and concentrations isolated from cucumber
The encumbrance of 62 populations of RKN also varied with PP-3 at two different temperatures (ambient and 30 ± 2°C) and concentrations (104 and 105 spores per ml). The encumbrance level of PP-3 was maximum at 30 ± 2°C than ambient temperature and spore concentration was also directly related to attachment with nematode cuticle (Table 5). The populations selected on the basis of maximum attachment with PP-3 were FSD/C4 (Faisalabad), JNG/C2 (Jhang), KHW/C9 (Khanewal), MUL/C2 (Multan) and RWP/C10 (Rawalpindi).
Encumbrance of PP-J with different RKN populations at different temperatures and concentrations isolated from cucumber
A great variability of the number of spores attached to the cuticles of J2 was observed when same 62 populations of RKN were evaluated for encumbrance with PP-J at different temperatures and concentration (Table 5). Two populations showed maximum encumbrance with PP-3 and PP-J were JNG/C2 (Jhang) and MUL/C2 (Multan) while other three populations having maximum encumbrance were FSD/C12 (Faisalabad), KHW/C3 (Khanewal) and RWP/C2 (Rawalpindi). The two RKN populations from cucumber which produced least encumbrance throughout the in-vitro were RWP/C5 (Rawalpindi) and RWP/C6 (Rawalpindi).
Assessment of genetic variability among different populations of root knot nematodes using PCR-RFLP of CO II and large subunit of rRNA gene (lrRNA) of the mitochondrial genome
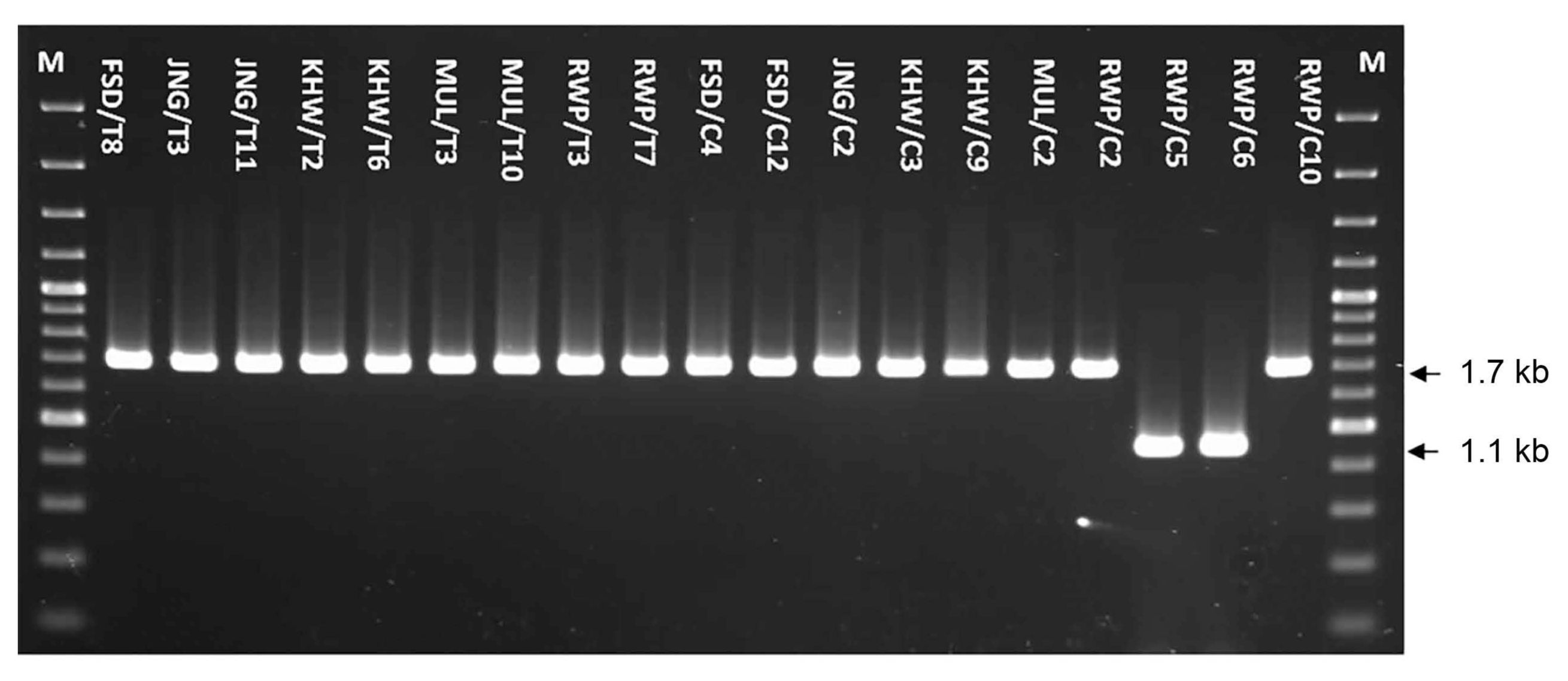
The primer set used to amplify intergenic region between CO II and large subunit of rRNA gene (lrRNA) of the mitochondrial genome. It produced single amplicon from each of the 19 samples (Fig. 3). A PCR product of 1.1 kb, expected from M. arenaria species, was obtained from the samples RWP/C5 and RWP/C6 which were chosen on the basis of minimum encumbrance, hence these samples were confirmed as M. arenaria. The seventeen samples, selected on the basis of maximum encumbrance, amplified a product of 1.7 kb related in size expected from M. incognita and M. javanica species (Fig. 3).
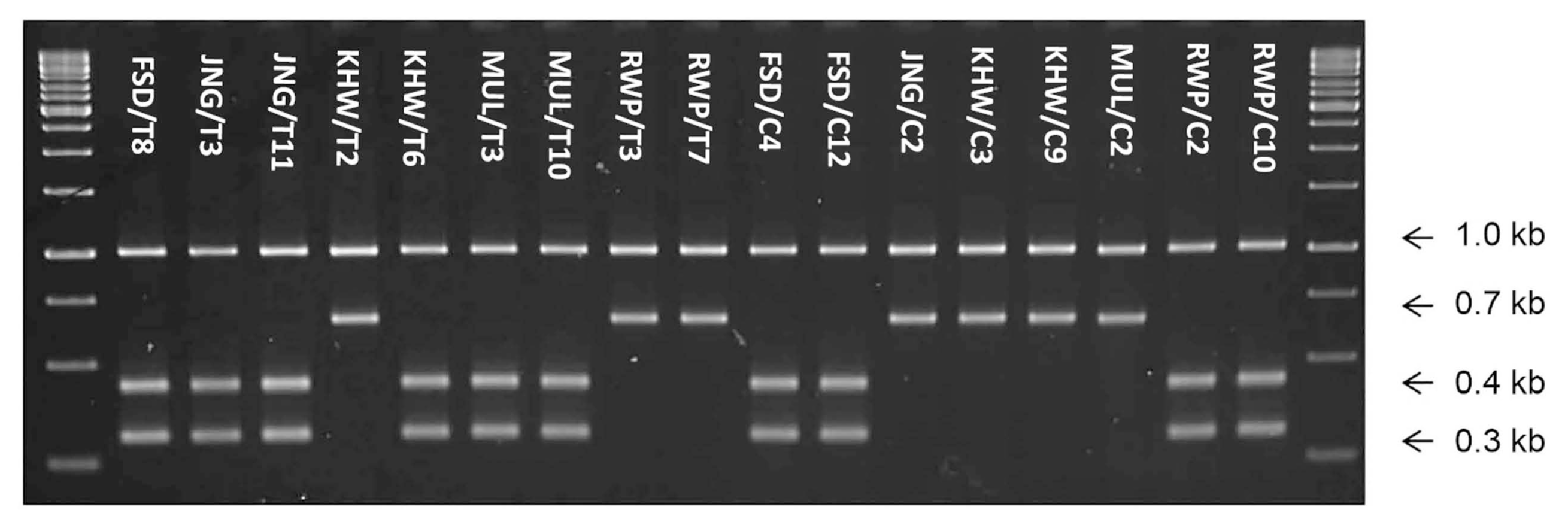
The Hinf I restriction analysis of 1.7 kb fragment to differentiate among Meloidogyne species determined single digestion site in samples KHW/T2, RWP/T3, RWP/T7, JNG/C2, KHW/C3, KHW/C9 and MUL/C2, generating two fragments of 1.0 and 0.7 kb (Fig. 4).
These restriction fingerprints related the samples to M. javanica. In rest of 10 samples including FSD/T8, JNG/T3, JNG/T11, KHW/T6, MUL/T3, MUL/T10, FSD/C4, FSD/C12, RWP/C2 and RWP/C10, an additional enzyme digestion site cleaved the 0.7 kb fragment to generate two more fragments of about 0.4 and 0.3 kb (Fig. 4). This restriction pattern related the samples to M. incognita species. In conclusion, M. incognita was found most prevalent in the 19 samples with the highest frequency (52.6%) followed by M. javanica (36.8%) and M. arenaria (10.5%).
Discussion
Encumbrance of Pasteuria endopsores to the cuticle plays an important role in biological management of root knot nematodes (Channer and Gowen, 1992; Espanol et al., 1997; Stirling, 1984; Stirling et al., 1986; Vagelas et al., 2012; Wishart et al., 2004). Different populations of Pasteuria endospores exhibit different levels of encumbrance to nematode cuticle. These variations have been attributed to differences in the surface composition of different species, races and RKN populations (Davies and Danks, 1992; Davies et al., 2008) and to the heterogenity of the P. penetrans endospore surfaces themselves (Davies et al., 1992). The specificity of endospore encumbrance to J2 cuticle has been studied biochemical and immunological methods. The results showed a high degree of heterogeneity both within and among different populations of P. penetrans (Davies and Redden, 1997; Preston et al., 2003). A carbohydrate-protein mechanism is involved in endospore encumbrance to M. incognita (Davies and Danks, 1993; Persidis et al., 1991) while according to Spiegel et al. (1996) carbohydrate residues, carbohydrate-recognition domains and a 250-kDa antigen on the J2 cuticle of M. javanica were shown to be involved in Pasteuria endospore attachment. According to Davies and Opperman (2006) collagen like proteins on Pasteuria spore core are involved for the adhesion of the endospore to J2 cuticle. These differences in attachment indicated complex interactions between the cuticle of the nematode and the surface of the Pasteuria endospore (Tian et al., 2007).
In current work interspecific and intraspecfic variability in encumbrance was observed with two Pasteuria isolates at different temperatures and concentrations. Temperature is one of several environmental factors that influence the ability of endospore to adhere and to infect Meloidogyne J2 (Hatz and Dickson, 1992). The rate of spore encumbrance to J2 increased with increasing temperature and current results are similar to previous works (Giannakou and Gowen, 2004; Singh and Dhawan, 1990; Stirling et al., 1990; Zareen et al., 2002). Pasteuria penetrans spores attached more readily at 22.5-30°C than 15°C (Stirling, 1981). However, Giannakou et al. (1997) observed greater attachment at 25 and 30°C but the maximum number of endospores was attached at 30°C (Ahmed, 1990; Hatz and Dickson, 1992; Orui, 1997). At higher temperature (above 30°C) the rate of encumbrance with J2 declined (Hatz and Dickson, 1992). Exposing nematodes to 30°C also increased rate of encumbrance. Freitas et al. (1997) pre-exposed M. arenaria J2 in 30°C water before exposure to endospores increased J2 receptivity to endospores when compared to treatments at 25°C and 35°C. The variable encumbrance at different temperatures is due to the fact that it is dependent on nematodes’ mobility. At 30°C the mobility of nematodes is high while above 35°C or below 5°C nematodes have little activity (Taylor and Sasser, 1978). That’s why at these temperatures attachment is decreased. Other reason is that, the bacterium developed more quickly within its host at 30°C and 35°C than at 25°C or below (Hatz and Dickson, 1992).
Encumbrance of Pasteuria endopsores to the J2 cuticle is also dependent on Pasteuria spore densities/concentrations. As the Pasteuria endospores are non-motile and successful encumbrance with J2 cuticle is only achieved when a suitable nematode comes under its vicinity (Chen and Dickson, 1998). The rate of encumbrance to J2 increased approximately five to six times for each 10-fold increase in numbers of Pasteuria endospores in the suspension Hewlett and Dickson (1993). Zareen et al. (2002) reported that encumbrance of Pasteuria endospore to J2 cuticle reduced with increase in dilution factor of spore suspension while Alves et al. (2004) provoked that in order to improve the endospore attachment, endospore concentration of the suspension is more important than to increase the agitation period of the nematodes in the bacterial suspension. Pasteuria endospore concentration in soil is also key factor for its infectivity (Stirling et al., 1990). Endospore densities in soil was positively correlated with percentage of J2 attached and number of spores/J2 (Talavera and Mizukubo, 2003).
The PCR reaction amplified intergenic region between cytochrome oxidase subunit II gene (COII) and large subunit of rRNA gene (lrRNA) of the mitochondrial genome. The primer C2F3 and 1108 identified M. incognita with the highest frequency (52.6%) followed by M. javanica (36.8%) and M. arenaria (10.5%). Heterogeneity in band size was detected in mitochondrial genome of different Meloidogyne species. The sizes of PCR products were 1.7 kb for M. incognita and M. javanica populations while populations of M. arenaria produced 1.1 kb fragment. Therefore, M. arenaria was easily distinguished from M. incognita and M. javanica based on band size of amplified PCR products. No band size variability was detected within species of Meloidogyne. The digestion with Hinf I yielded three different fragment length patterns on 1.5% agarose gel. First, restriction digestion of 1.7 kb M. javanica amplification product determined one enzyme digestion site and resulted in two fragments of 1.0 and 0.7 kb. Second, an additional enzyme digestion site on M. incognita product cleavage the 0.7 kb fragment to generate two more fragments of about 0.4 and 0.3 kb. Third, M. arenaria had no enzyme digestion site by Hinf I digestion.
For precise identification of Meloidogyne species, mitochondrial genome provides good source of genetic markers for identification (Blok et al., 2002; Hu and Glasser, 2006; Hugall et al., 1994; Jeyaprakash et al., 2006). Intraspecfic mitochondrial variation has been observed in a region of multiple nucleotide repeating units in Meloidogyne. Mitochondrial DNA has been applied not only for the identification of Meloidogyne species but also for the molecular differentiation or population genetic study of nematodes (Liu et al., 1999; Szalanski et al., 2000). In the present study intergenic region between cytochrome oxidase subunit II and 16S ribosomal mitochondrial genes have been amplified to determine intra-Meloidogyne genetic variability. The assay successfully differentiated isolates in to three types i.e. M. incognita, M. javanica and M. arenaria based on variation in fragment sizes of the PCR products/their restriction fingerprints. The similar studies have been conducted on characterization of USA (Powers and Harris, 1993) and Korean isolates (Han et al., 2004; Oh et al., 2009) of Meloidogyne species exploiting the same genetic markers. For M. arenaria C2F3/1108 amplified PCR product was of 1.1 kb in USA isolate (Powers and Harris, 1993; Powers et al., 2005) which is of the same size as found in current studies on Pakistani M. arenaria isolates. However, Oh et al. (2009) reported 1.7 kb fragment amplified from Korean M. arenaria isolates. PCR amplification product of Pakistani M. javanica isolates exhibited 1.7 kb product size and on digestion resulted into two band sizes of 1.0 and 0.7 kb which was similar to previous work of Powers and Harris, 1993. However, Hinf I digestion revealed results differing from those of previous study (Oh et al., 2009). Therefore M. incognita, M. javanica and M. arenaria has significant nucleotide variations depending upon different ecological origins.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






