

Glomerella leaf spot (GLS) is a relatively recent and destructive disease of apple, but still restricted to a few world regions. GLS was reported for the first time in the Southwest of USA in 1970 (Taylor, 1971) but only at the end of the 90s became more important when a severe outbreak was reported (Gonzalez and Sutton, 1999). In 1983, GLS appeared in Southern Brazil, and since then it has been considered one of the major diseases of apple (Sutton and Sanhueza, 1998). In 2012, it was reported in Eastern China as a new destructive disease of apple (Wang et al., 2012). Later on, in 2016 it was noticed in Uruguay causing severe defoliation in a commercial apple orchard located in the Southern part of the country (Casanova et al., 2017).
The symptoms of GLS appear during the summer on leaves and fruits of apple. On leaves they consist in brown to ochre necrotic spot lesions that grow quickly taking an irregular shape and causing generalized chlorosis. Under warm and hot conditions, severe defoliation of the affected tree can be observed. On fruits, circular, slightly sunken and corky lesions of 0.5 to 3 mm in diameter and 0.5 to 2 mm in deep appear; these lesions are usually surrounded by a red border (Casanova et al., 2017; González and Sutton, 1999, 2004; Wang et al., 2012). Cultivars as Golden Delicious and their descendants such as cv. Galaxy or Cripps pink are highly susceptible, but others like Red Delicious and Fuji are highly resistant (Liu et al., 2016; Velho et al., 2015; Wang et al., 2015b). In Uruguay, commercial apple production is concentrated in the South where 22% of orchard trees belong to cvs. Galaxy and Cripps pink (MGAPDIEA, 2016).
Phenotypic analyses initially identified Glomerella cingulata, Colletotrichum gloeosporioides and C. acutatum as the causal agents of GLS in Brazil and USA (González and Sutton, 1999, 2004; Sutton and Sanhueza, 1998) and to G. cingulata in China (Wang et al., 2012). Later, using multilocus genetic markers the species associated to GLS in Brazil were reidentified as C. fructicola (Velho et al., 2015) and C. karstii (Velho et al., 2014) and C. fructicola and C. aegnima in China (Wang et al., 2015b).
GLS has gradually spread to new apple production regions, increasing its economic impact worldwide. However, partially due to its recent emergence, only a few studies have been carried out so far, particularly those focusing on epidemiology and control. Acervuli and perithecia containing conidia and ascospores are produced in lesions of infected leaves (Casanova et al., 2017; Valdebenito-Sanuheza et al., 2002), but the role of both types of spores in the development of the disease, is poorly understood. Pathogenicity tests with leaves and fruits by means of artificial inoculation with conidial suspensions of Colletotrichum spp. have been successfully conducted (Casanova et al., 2017; Velho et al., 2014, 2015; Wang et al., 2015b). In contrast, the ability of ascospores in producing symptoms of GLS on both leaves and fruits has not been demonstrated.
In order to better understand the disease occurrence in Uruguay, the objectives of our study were i) monitor the distribution and evolution of GLS symptoms on commercial apple orchards ii) identify Colletotrichum species involved with GLS symptoms in fruit and leaves, and iii) test the ability of ascospores in causing GLS symptoms on apple leaves and fruits.
Materials and Methods
GLS field symptoms and fungal isolates
In summer 2015 and 2016, the presence of symptoms and their distribution were observed in trees of the GLS-diseasedorchards composed of cv. Cripps pink with cv. Galaxy as pollinator in Kiyu, San José Department (34° 38′*S* 56° 45′*W*), Uruguay. Leaves and fruits with typical GLS symptoms were collected from trees randomly selected; a maximum of three samples per tree including fruits and leaves were taken.
To isolate the fungus from the lesions, the fruits were first disinfected with 70% ethanol. After drying, fruit peel was removed from the affected area with a sterile scalpel leaving the internal lesion exposed. Small portions of the corky lesion were plated in potato dextrose agar (PDA) (Oxoid Ltd., Hampshire, England) and incubated at 25°C in the dark for 5 days. Hyphae growing out from fruit tissues were subcultured onto PDA media.
Monoascospore and monoconidial isolates were obtained directly from leaf lesions under magnifying glass. For this, a single perithecium or acervulus was transferred from leaf lesions to a tube containing 300 μl of sterile distilled water (SDW) (one per leaf) and vigorously stirred using a vortex. Immediately, 100 μl of ascospores or conidial suspension was placed onto PDA supplemented with 0.4 g/l of streptomycin sulphate (Sigma-Aldrich, China). After 24 h of incubation at 25°C in the dark, a germinated ascospore or conidium was transferred to PDA medium and incubated to the same conditions.
The Colletotrichum spp. isolates were stored at −20°C in colonized sterile paper filters dried with silica-gel (Peever et al., 1999). Isolates were deposited in the culture collection of the Department of Plant Protection, Faculty of Agronomy, University of the Republic Uruguay.
Morphological characterization
All Colletotrichum isolates were grown on PDA medium for two to four weeks at 25°C and 12 h photoperiod under near ultraviolet light. Macroscopic characteristics, such as colony colour and presence of reproductive structures as perithecia, acervuli, asci, ascospores and conidia were recorded.
PCR amplification, sequencing and phylogenetic analysis
Mycelium and reproductive structures from all isolates grown on PDA for one week at 25°C in the dark were scraped off with a spatula and placed in 2.0 ml microtubes. Total DNA was extracted using a protocol described by Paolocci et al. (1999).
The partial actin (ACT), β-tubulin (BTUB2) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene regions were amplified and sequenced for all Colletotrichum sp. isolates. The primer pairs used were ACT-512F/ACT-783R (Carbone and Kohn, 1999), BT2Fd/BT4R (Woudenberg et al., 2009) and GDF1/GDR1 (Guerber et al., 2003) respectively.
Each PCR reaction contained 1x PCR buffer, 2.5 mM MgCl2, 400 μM of each dNTP, 0.4 μM of each primer, 1 U of DNA polymerase (Tiangen, China) and 1 μl of template DNA. The PCR reaction mix was adjusted to a final volume of 40 μl with MQ water. The amplifications were performed on a MultiGene™ Mini (Labnet International, Inc., USA). The program run was as follows: an initial step of 94°C for 5 min followed by 34 cycles of denaturation at 94°C for 45 s, annealing at 52°C (57°C for ITS) for 30 s, and elongation at 72°C for 45 s. A final extension was performed at 72°C for 10 min. The PCR products were analyzed in 1.5% agarose gels stained with GelRed™ and visualized under UV light. A gene ruler 100-bp DNA ladder plus was used as a molecular weight marker (Thermo, Lithuania).
The PCR products were purified and sequenced in Macrogen Inc., Seoul, Korea. The DNA sequences were edited using DNAman 8.0 program (Lynnon Corporation). The sequences were aligned using ClustalW program (Larkin et al., 2007), available within MEGA 7.0.26 program (Kumar et al., 2016). The sequences derived from this study were deposited in the GenBank (Table 1). In the alignments, related sequences and sequences of the phylogenetically closest species (including ex-type) of C. boninense and C. gloeosporioides species complexes obtained from the Gen-Bank, were incorporated (Damm et al., 2012; Weir et al., 2012). The ex-type of C. acutatum CBS 112996 was used as the out-group. The sequence alignment was deposited in TreeBASE (www.treebase.org) under the accession number S21955.
Phylogenetic analyses were performed for each gene region separately as well as for the multi-locus alignment using MEGA 7.0.26 program for maximum likelihood (ML) and maximum parsimony (MP) analysis. To confirm that the three sequence datasets were congruent and combinable, highly supported clades among the trees generated from the different data sets for MP and ML analysis were compared to detect conflicts. High support typically refers to bootstrap support values ≥ 70% (Hillis and Bull, 1993).
ML analysis was based on the Tamura-Nei model and the tree was drawn to scale, with branch lengths measured in the number of substitutions per site. MP analysis was performed using the subtree pruning regrafting (SPR) algorithm with search level 1, in which the initial trees were obtained with random additions of sequences (10 replicates). All characters were unordered and of equal weight. Gaps were treated as missing data. The tree length (TL), consistency index (CI), and retention index (RI) were recorded. The robustness of the the ML and MP trees was assessed by bootstrap analysis with 1,000 replicates (Hillis and Bull, 1993).
Pathogenicity tests
The pathogenicity of the isolate F51of C. karstii was tested with conidia suspension, whereas the perithecial isolates L49, L53, L64, F71 and F84 of C. fructiola were used to verify the ability of ascospores in causing GLS symptoms. The six strains were randomly selected and tested against both apple fruits and leaves.
Isolates were grown on PDA at 25°C for three weeks with a 12 h photoperiod under near ultra violet light to induce sporulation. To prepare ascospore suspensions, perithecia were removed with sterile fine forceps from the colonies of C. fructicola growing on PDA, placed into sterile tube of 2 ml with SDW and shortly vortexed during one minute. To prepare the conidial suspension, colony of C. karstii growing on PDA was flooded with 10 ml of SDW and then scraped with a sterile spatule. In both cases, the resulting suspension was filtered through two layers of cheese cloth and adjusted with a counting chamber to 1 × 105 ascospores or conidia/ml.
Young terminal twigs of cv. Galaxy were collected from a disease-free commercial apple orchard. For the first assay, fully developed young leaves were selected and placed individually in Petri dishes containing moistened sterile paper filter. To avoid the dehydration, the petiole of each leaf was wrapped with wet cotton. The detached leaves were inoculated by spraying (three leaves per isolate) and then incubated at 25°C. Control leaves were spraying with SDW.
For the second assay, another set of young terminal twigs with 5-6 leaves were planted individually in glass flasks containing wet sand. The twigs were inoculated by spraying the spore suspension (five twigs per isolate). They were immediately covered with a moistened plastic bag and incubated at 25°C. Control twigs were spraying with SDW.
Asymptomatic apple fruits of the cv. Cripps pink at the beginning of colour change were harvested from a disease-free commercial orchard. All fruits were surface-disinfested with 70% ethanol. After that, fruits were inoculated by spraying (five per treatment) and incubated in moistened plastic bags at 25°C. Control fruits were spraying with SDW.
All assays were monitored daily; the initial symptoms and their evolution as well the presence of reproductive structures of Colletotrichum were recorded. C. karstii was reisolated from leaves and fruits to complete Koch’s postulates.
Results
GLS field symptoms and fungal isolates
Typical GLS symptoms (Casanova et al., 2017) were observed on leaves and fruits in both years and on both apple cvs., Cripps pink and Galaxy. The first GLS disease symptoms were observed during January, primarily occurring in leaves (Fig. 1A). The leaf lesions rapidly evolved into chlorosis (Fig. 1B) and, approximately one month later, a severe defoliation (approximately 50% of fallen leaves) was observed (Fig. 1E and 1F). The fruit lesions did not evolve into rot remaining as small corky lesions (Fig. 1C).
The spatial distribution of symptoms was widespread randomized within orchards as well as individual trees. In each apple tree, infected leaves and fruits were observed at different heights, even in the lower inner tree canopy and terminal twigs (Fig. 1D). Presence of acervuli (Fig. 1H) and perithecia (Fig. 1I) containing conidia and ascospores, respectively, were consistently observed in leaf lesions (Fig. 1G), but never in fruit lesions.
Typical colonies of the Colletotrichum fungus, i.e. showing white to orange or white to gray mycelium colour, presence of orange-salmon gelatinous masses composed of hyaline, fusiform to obtuse conidia, successfully grew from fruit and leaf infected tissues. Forty-six isolates of Colletotrichum were obtained, 21 originated from leaves and 25 from fruits (Table 1).
Morphological characterization
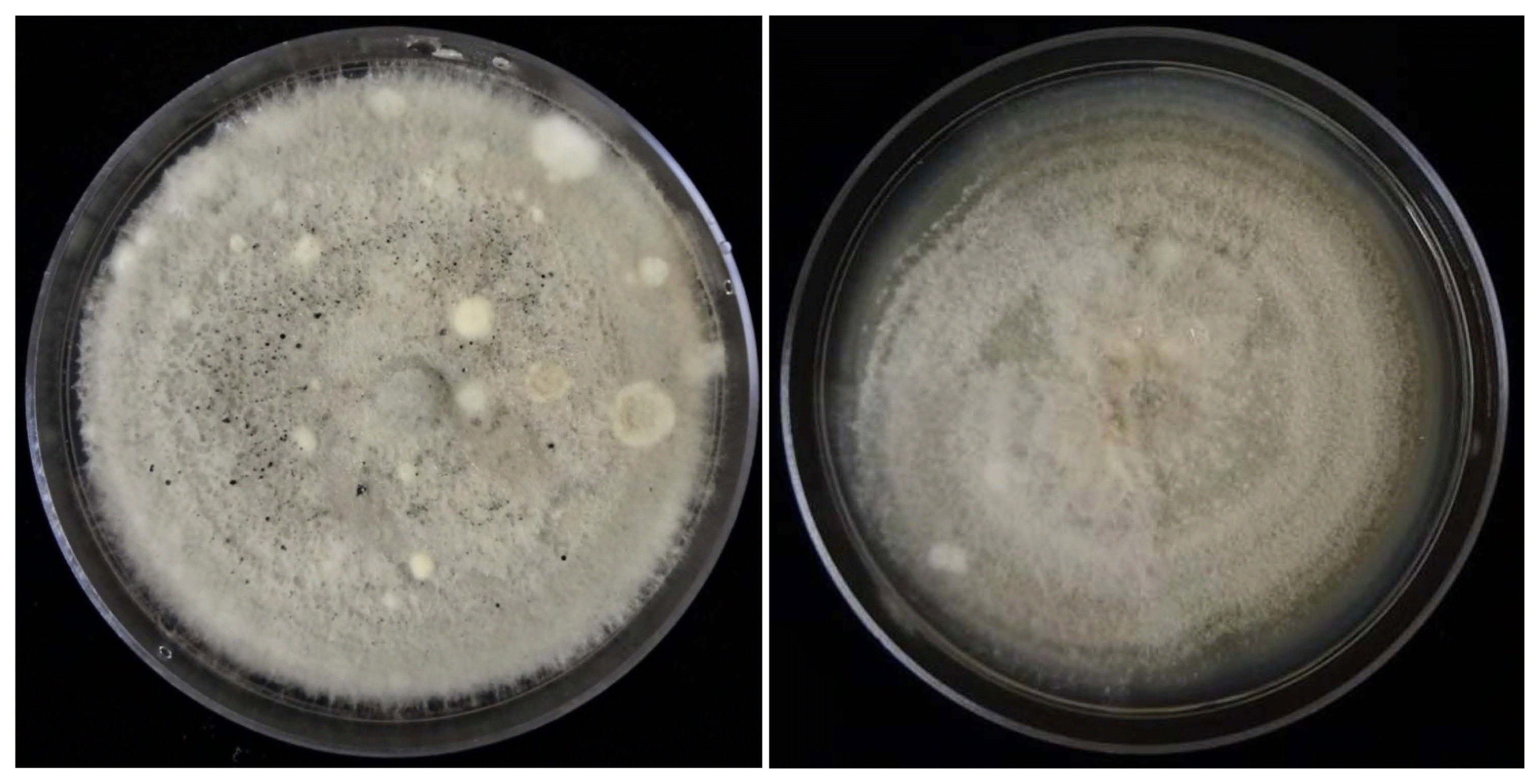
Initially the colony color of all the isolates on PDA was predominately white, but a few days later, they gradually changed to pale-gray to gray.
Reproductive structures were observed in all colonies. For the strains phylogenticaly identified as C. fructicola, the predominating structures were perithecia containing asci with ascospores, while for those of C. karstii only conidia were found (Fig. 2).
PCR amplification, sequencing and phylogenetic analysis
Amplicons of approximately 260, 500 and 250 bases were amplified for the ACT, BTUB2 and GAPDH genes regions, respectively, in all isolates (Table 1).
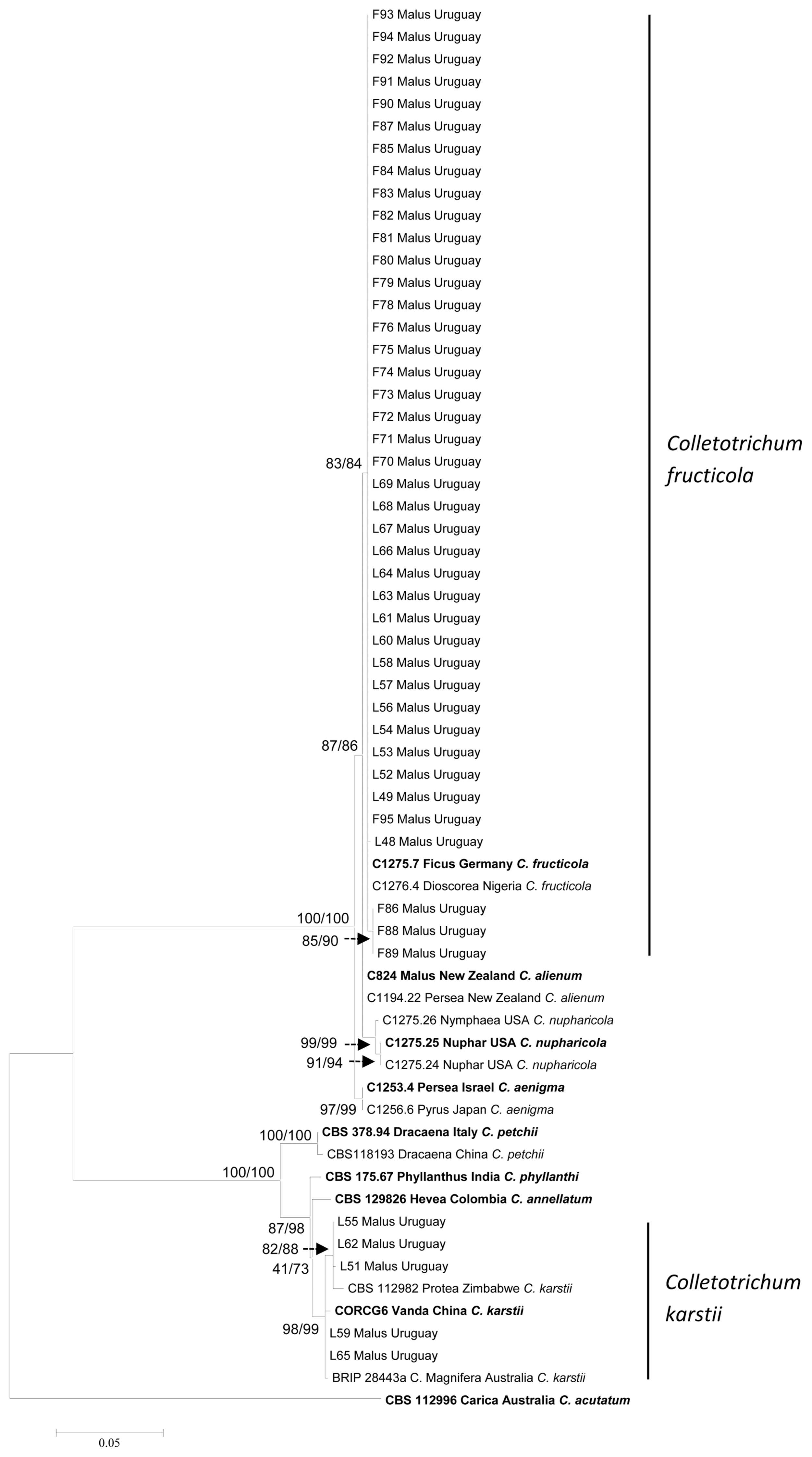
The three sequence data sets of species belonging C. boninense and C. gloeosporiodes species complex, did not show major conflicts in tree topology, indicating that the three genes could be combined. The multi-gene analysis consisted of 905 characters (including alignment gaps), of which 216 characters were parsimony-informative. MP analysis resulted in three most parsimonious trees (TL = 369 step, CI = 0.94, RI = 0.99).
Two main clades are recognized in the phylogenetic tree drawn from the multi-gene analysis. One clade is composed of species included in the C. gloeosporioides species complex and most of the Uruguayan isolates resolved as C. fructicola (41 isolates). The other main clade is composed of species included in the C. boninense species complex and five Uruguayan isolates identified as C. karstii. The main clades and sub-clades bootstrap support, robustly resolved all Colletotrichum species for both methods of analysis (Fig. 3). The isolates identified as C. fructicola belong to the same haplotype, except L48 that differs in one nucleotide in the BTUB2 region. With regard to C. karstii three haplotypes were found, L55 and L62, L59 and L65 with two different nucleotides in the GAPDH region and L51 with one different nucleotide in the BTUB2 region.
Pathogenicity tests
Both conidia of C. karstii and ascospores of C. fruticola, caused typical GLS symptoms in apple leaves and fruits (Table 2). No symptoms were observed in control treatments (Fig. 4A).
The first symptoms consisted in purple to ochre necrotic spot-like lesions on leaves and were already visible three and seven days after inoculation with C. fructicola (Fig. 4B and C) and C. karstii, respectively. Lesions in leaves inoculated with C. fructicola evolved rapidly to larger, irregular shape and necrotic lesions at seven days from inoculation (Fig. 4D and E) and to severe defoliation three days later (Fig. 4F). When inoculated with C. karstii the defoliation was mild at ten days from the inoculation (Fig. 4G). Abundant acervuli were present in leaf lesions caused by isolates of C. fructicola (Fig. 4D), but never were observed in leaf lesions caused by C. karstii.
On fruits, symptoms similarly appeared 21 days after inoculation when inoculated with C. fructicola or C. karstii isolates. The symptoms consist in small corky lesions that did not evolve to rot (Fig. 4H and I). No reproductive structures were observed on these lesions.
Cultural and conidial characteristics of isolates obtained from inoculated leaves and fruits with C. karstii, were confirmed on PDA as previously described fulfilling Koch’s postulates.
Discussion
In Uruguay, GLS was recently reported by Casanova et al. (2017) occurring in a small producing area and, to the best of our knowledge, it still remains restricted to the same locality. The reasons why GLS has not yet spread further are unknown. Microclimatic conditions in this locality seem particularly conductive to GLS development and may indeed have helped to create this situation. It is known that GLS requires warm temperatures and high humidity (Casanova et al., 2017; González and Sutton, 1999; Velho et al., 2015; Wang et al., 2015a). The GLS-infected apple orchards were established in a low area surrounded by windbreaks, hindering ventilation and possibly resulting in better environmental conditions to the pathogen. Furthermore, the affected area is slightly distant from other commercial apple orchards; being the nearest seven kilometers away.
The widespread randomized distribution of symptoms observed throughout trees and orchards where GLS is present, suggests that ascospores may play a pivotal role in the dispersion of this disease. In this work, ascospores were proofed for the first time to be pathogenic to leaves and fruits confirming its potential to cause infections. The frequent presence of acervuli and perithecia containing conidia and ascospores, respectively, in leaf lesions indicates that both types of spores are regularly produced by most strains of Colleotrichum causing GLS. In contrast to asexual spores of Colletotrichum which are rain-splash-dispersed (Perfect et al., 1999; Sutton and Shane, 1983), those sexually formed are wind-dispersed (Sutton and Shane, 1983). Conidia are released by rain splash and carried by water droplets to short distances generating a disease spatial gradient in focus pattern within the orchard. On the other hand, ascopores are ejected up to a small distance and, usually, dispersed by air currents reaching longer distances and leading to a randomized disease distribution throughout the orchard. The occurrence of symptoms in leaves on the top of the apple tree canopy, strongly suggests that infection has happened with ascospores carried by wind.
In this work it was confirmed that Colletotrichum remains alive in the small fruit corky lesions. Cheon et al. (2016) also were able to isolate C. gloeosporioides from symptoms in apple fruits comparable to those of GLS. Furthermore, this fungus did not result in rot development, probably because it could not grow out from small cork-like lesions to surrounding healthy tissues of fruits. Interestingly, in this confined situation it did not build any reproductive structure. It seems that apple fruits can active some resistance mechanisms able to avoid fungus growth from this lesion. Although most GLS-isolates do not have the ability to produce fruit rot, there are a few able to do this (Velho et al., 2015). Thus, investigating the reasons why GLS-isolates are in general not able to cause fruit rot would be an exciting topic for future research.
Based on phylogenetic analysis of GAPDH, BTUB and ACT genomic regions and pathogenicity test, C. karstii was found as a new species causing GLS in Uruguay. C. fructicola had already been reported associated with this disease by Casanova et al. (2017), but in this work appears as the dominant species (89%). C. fructicola and C. karstii were identified in Brazil as the causal agent of GLS (Velho et al., 2014, 2015) and C. fructicola as one of the two species associated to GLS in China (Wang et al., 2015b).
The results of this and other studies (Rockenbach et al., 2016; Velho et al., 2015) indicate that C. fructicola have an extremely aggressive behavior; within three days it causes visible symptoms on leaves inoculated with conidia or ascospores and a few days later severe defoliation. Moreover, under favorable conditions, warm temperatures and high relative humidity, abundant reproductive structures are formed on leaf lesions. In contrast, C. karstii develops much more slowly and reproductive structures seldom appear. Likewise, all isolates of C. fructicola grown in laboratory produced conidia and perithecia containing ascospores, but not C. karstii.
The species of Colletotrichum found in Uruguay causing GLS are similar to those reported in Southern Brazil. This suggests that apple plants imported from Brazil could have introduced the disease in Uruguay. Another hypothesis is that isolates of C. fructicola and C. karstii from Uruguay responsible for GLS have arisen from isolates capable of causing bitter rot. However, C. karstii has not been associated with bitter rot disease in Uruguay (Alaniz et al., 2015). Moreover, Uruguayan populations of C. fructicola causing bitter rot disease were found to be genetically distinct from Brazilian isolates obtained from GLS and bitter rot diseases (Rockenbach et al., 2016). Therefore, new population studies are needed to compare genetic structure of C. fructicola populations causing GLS and bitter rot in Uruguay and Brazil. This might help to elucidate the origin of the GLS in Uruguay.
In summary, the consistency and aggressiveness of the symptoms observed in this and other studies (Gonzalez and Sutton, 1999; Sutton and Sanhueza, 1998; Wang et al., 2015b), indicate the high impact potential of GLS in causing severe losses on susceptible cultivars if GLS spreads out to further apple producing areas in Uruguay. Developing new strategies for GLS management is extremely necessary because control methods have been currently based on regular applications of fungicides during the summer. Probably, the simultaneously production of ascospores and conidia might be, however, difficult due to their distinct and complementary roles in the development of disease epidemics.


 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






