

Phellodendron amurense is a deciduous tree of the family Rutaceaceae (Tang and Eisenbrand, 1992). Originating in northern China, South Korea and Japan, P. amurense has been cultivated in East Asia and Eastern Europe to produce bark for medicinal substances used as natural drugs, whereas the wood of this tree is used for furniture making and industrial art (Azad et al., 2005). This species is mainly distributed in Liaoning, Jilin, Heilongjiang and Inner Mongolia in China (Jiao et al., 2017). There is a long history of medicinal use of P. amurense, which has been listed in the Chinese Pharmacopoeia, with properties of reducing inflammation and preventing infection by pathogen fungi, as well as treating diarrhoea, jaundice, ulcers, lupus erythaematousus, eczema and other diseases (Tang and Eisenbrand, 1992). A recent study by Li et al. (2018) first reported Nectria haematococca to be a cause canker rot in P. amurense in a Huashan Forest Farm and Yangtianwo Drug Farm in Dayi County, Chengdu, Sichuan Province. This disease causes decaying of the trunk cortex, shedding of branches and wilting, affecting the yield of the medicinal materials, reducing the medicinal value and resulting in substantial loss.
At present, the main control method of plant diseases is the use of chemical pesticides. However, long-term use of chemical pesticides causes environmental pollution, and pesticide residues lead to an increase in disease resistance and reduced soil fertility (Passari et al., 2018). Moreover, chemical control causes the deterioration of natural resources, degradation of product quality, and destruction of ecological balance, with serious impacts on human health and ecological safety (Abbas et al., 2017). In contrast, biocontrol technology, which utilizes beneficial microorganisms to control the occurrence and development of disease, has been proposed as a solution (Iasur-Kruh et al., 2018). Indeed, the development of effective biological agents against pathogenic fungi is an important method to controlling plant diseases (Boukaew et al., 2017). Biocontrol bacteria are ubiquitous in the environment, with a wide range of prevention and control properties; furthermore, such bacteria exhibit good effects in controlling plant diseases, and there is no pathogen resistance. Thus, biocontrol bacteria are highly promising for plant disease control (Boukaew et al., 2017; Etesami and Alikhani, 2017; Kamal et al., 2016). Biocontrol microbes are abundant and include bacteria, fungi and actinomycetes (Baltz, 2016). The relevant bacteria are largely Pasteurella spp., Pseudomonas spp., Agrobacterium radiobacter and Bacillus spp. Among them, Bacillus spp. are known for their ability to control plant diseases through various mechanisms, including the production of secondary metabolites. Bacillus species, especially B. subtilis, B. cereus and Lactobacillus plantarum, are effective against plant diseases caused by soil transmission as well as leaf and post-harvest fungal pathogens (Yu et al., 2011). Most bacteria of the genus Bacillus are heat resistant, drought resistant and ultraviolet irradiation resistant (Obagwu et al., 2003). Almost all healthy plants contain endophytic bacteria, which colonize intercellular spaces and have many effects on host plants, such as promoting growth, preventing disease and fixing nitrogen. Accordingly, these bacteria are a natural resource for the biological control of plant diseases, with broad research value and development prospects. Some researchers have isolated antagonistic endophytic bacteria with good control effects against many diseases from a variety of plants, such as cotton, potato and pepper (Faltin et al., 2004).
To date, there have been few reports on canker rot in P. amurense in China and worldwide. Only Li et al. (2018) isolated and identified a causative pathogen: N. haematococca, whereas research on the screening of antagonistic bacteria from P. amurense and their application against canker rot has not yet been reported. In this study, endophytic bacteria were isolated and screened from healthy tissues of P. amurense from a forest farm and then assessed for their ability to effectively inhibit the pathogenic fungus responsible for canker rot in P. amurense. Morphological, physiological, biochemical and molecular characterizations were performed. The control effect was evaluated in pot and field experiments. The antimicrobial activity and mechanism of antagonistic strains were determined, and the effects of antagonistic strains on mycelial morphology and spore germination were determined. The aim of this study is to provide support for the development of new biocontrol agents for canker rot in P. amurense and to provide a more economical, efficient and environmentally friendly solution for prevention and control of this disease (Kumar and Hwang, 2006).
Materials and Methods
Materials. Plant samples and disease fungus
One-yearold healthy P. amurense seedlings were used. Healthy bark of P. amurense was collected from Huashan Forest Farm (Elevation 1248 m, 30°37′01.1″N, 103°18′46.1″E) and Yangtianwo Drug Farm (Elevation 1260 m, 30°37′06.9″N, 103°19′18.3″E) in Dayi County, Chengdu, Sichuan Province, China.
Fungus
The pathogenic fungus N. haematococca isolated from P. amurense canker rot was provided by the Key Laboratory of Forest Protection of Sichuan Province (Li et al., 2018).
Culture medium: (1) PDA: potato 200 g, glucose 20 g, agar 15-20 g, distilled water 1000 ml, pH 7.0. Potato glucose agar was used for pathogen culture. (2) NA: peptone 10 g, beef extract 3 g, sodium chloride 5 g, agar 15-20 g, distilled water 1000 ml, pH 7.4-7.6. Beef peptone solid medium was used for the isolation and purification of bacteria.
Isolation of biocontrol endophytic bacteria
To isolate endophytic bacteria, the bark of P. amurense was disinfected with 70% alcohol and 3% sodium hypochlorite. The bark tissues were then washed several times with sterile water. The next step was to harvest the endophytic bacteria by the dilution separation method (Hornby, 1969). The thoroughly sterilized tissue was fully ground and diluted, and 10−3, 10−4, and 10−5 dilutions of 100 μl of the tissue solution were spread onto NA medium, and cultured for 1 d in a 3°C incubator to isolate single colonies distinctly different due to colony morphology.
Screening of antagonistic bacteria
According to the antagonistic bacterium screening method of Edwards and Mcbride (1975), antagonistic bacteria were preliminarily screened by the confrontation culture method. Strains with an obvious inhibition circle were selected, and their widths were measured. After purification, the strains were screened again. LB culture medium was selected for culture of antagonistic strains in a shaking bed incubator set at a constant temperature of 25°C and 135 r/min for 2 d. The fermentation culture filtrate was collected and filtered through filter paper and centrifuged at 4,000 r/min for 8 min, and the upper layer was filtered through a 0.22 μm membrane to prepare a sterile culture filtrate. The growth rate method of Bekierkunst and Szulga (1954) was used to determine strain antagonism. LB plates spread with sterile water and no fermentation culture filtrate were used as a control. Samples were cultured at 25°C constant temperature, and the process was repeated three times.
Identification of antagonistic bacteria
Morphologic observation: The antagonistic bacterium was inoculated onto NA medium and cultured in an incubator at 25°C. Morphological characteristics of colonies were assessed according to the Manual of Identification of the Common Bacterial System (Montanari et al., 2004; Zou et al., 2015).
Physiological and biochemical testing: The antagonistic bacterium was tested for Gram staining and spore staining as well as for the gelatine liquefaction test, nitrate reduction reaction, starch hydrolysis test, sugar oxidation fermentation test, contact enzyme reaction, acetyl methyl methanol test and hydrogen sulfide test.
Molecular biological identification: The strain was cultured in 20 ml of NA liquid medium for 24 h under constant temperature shock. The culture liquid was collected into 2 ml centrifuge tubes and centrifuged at 4°C and 12,000 rpm for 10 min. DNA was extracted according to the bacterial genome DNA kit (Tiangen Biochemistry Technology Company of Limited Liability). The extracted DNA was amplified by Polymerase Chain Reaction using the bacterial 16S rDNA universal primer (27F5’-AGAGTTTGATCCTGGCTCAG-3’) and (1492R5’-GGTTACCTTGTTACGAC -T T-3’).
Each 25 μl PCR mixture contained 0.5 μl DNA template, 12.5 μl 2 × Taq Master Mix, 1 μl each upstream and downstream primers, and 10 μl ddH2O. The reaction consisted of a 4-min pre-denaturation at 94°C, 35 cycles of denaturation 40 s at 94°C, annealing 30 s at 55°C, and extension for 1 min at 72°C, and a 10 min final elongation at 72°C. Five microlitres of the amplified product was assessed by 1% agarose gel electrophoresis and sequenced. The sequences were analysed by Blast homologous alignment, and the 16S rDNA sequences of highly homologous strains were downloaded from GenBank. Finally, a phylogenetic tree was constructed using the MEGA5.0 software adjacency method, and the phylogenetic status of the antagonistic strain was determined.
Determination of control effects in a pot experiment
The antagonistic bacterium was incubated for 48 h in NA liquid medium in a shaking bed incubator set at 28°C and 180 r/min. The culture medium was prepared with different concentrations of 104, 106, and 108 cfu/ml. Five pieces of pathogenic fungus mycelium (Φ=8 mm) were obtained by the punch method and added to each bottle of PDA liquid medium; pathogen spores were at a concentration of 105/ml after 72 h. One hundred healthy annual P. amurense seedlings were selected and divided into three groups, which were treated as follows: (1) plants were inoculated by wound inoculation with 20 ml of pathogenic fungus suspension, and after 15 d, 20 ml of antibiotic suspension at different concentrations was inoculated at the original site of inoculation by spraying (Sørensen et al., 2016); (2) plants were first treated with 20 ml of antibiotic suspension at different concentrations for 15 d and then inoculated with 20 ml of pathogen suspension; (3) plants were inoculated with different concentrations of antibiotic suspensions and 20 ml pathogenic fungus suspension at the same time. Sterile water served as the control. Each treatment was repeated ten times. The disease was investigated with aseptic water and aseptic culture medium for 30 d. The incidence, disease index and control effect were calculated. Disease was scored as follows: (I) no disease observed on the plant; (II) withering twigs less than 25%; (III) withering twigs 25%-50%; (IV) withering twigs 51%-75%; and (V) withering twigs more than 75%.
Field control
Field-control trials were conducted in a pure forest of P. amurense in Xieyuan Town, Dayi County, Chengdu City, Sichuan Province from July to August 2016. Two-year-old healthy P. amurense plants with similar growth potential were selected. A pathogen solution with a spore concentration of 107 cfu/ml and culture filtrate with 108 cfu/ml antagonistic bacterium were prepared. Concentrations of antibiotic solutions were 108, 2 × 106, 106, 5 × 105, 2.5 × 105, and 1.25 × 105, respectively. The following three types of processing were performed. (1) The pathogen was inoculated into a wound in the middle of a branch. At 15 days after inoculation, the undiluted solution of antagonistic bacterium and the diluted bacterial solution were inoculated at the same site using the spraying method (Sørensen et al., 2016). (2) The antagonistic bacterium was inoculated in 15 d in advance by spraying the antagonistic bacterium and each bacterial dilution, after which the pathogen solution was inoculated at the original site. (3) Inoculation with the pathogen solution, original solution and diluted solution was performed at the same time. Using 30 plants per treatment, each pathogen and antagonist was inoculated in 100 ml. Sterile water and culture medium were used as controls, and field management was carried out according to standard practices. Disease development was evaluated at 50 d after treatment. The plants were rated for disease to determine the control effect in potted plants.
Determination of the antagonistic mechanism of the antagonistic strain
Inhibitory effect of the antagonistic bacterium on the pathogenic fungus: (1) Effect on the mycelial growth of the pathogenic fungus - an antagonism test was carried out using the perforating method; sterile water was used as the control, and samples were assessed at 24, 48, 72, 96 and 120 h after addition of the antagonist supernatant. This assay was used to determine the diameter of the fungal colony and the diameter of the control fungal colony; each treatment was repeated 3 times. A growth curve of the disease after confrontation was drawn. After the control mycelium was covered with a flat plate, the difference between the mycelia of the surrounding colony and the normal mycelium of the control colony was observed under a light microscope (Hoyt et al., 2015). (2) Effects on the germination and production of spores by the pathogenic fungus: 107 cfu/ml of spore suspension was prepared, and a sterile filtrate was obtained. A conidial suspension of 2 μl of the pathogenic fungus was placed onto aseptic slides, and aseptic filtrates with an equal volume of concentrations of 1%, 5%, 25% and 50% were added to the conidial suspension droplets. Sterile water was used as the control, and the treatment was repeated 3 times. Conidial germination was detected at 5 h, 12 h and 24 h under the condition of moisture retention at 25%. Germination of the spore bud tube was considered at more than 1/2 of the spore diameter. (Sørensen et al., 2003). For each treatment, 200 spores were observed under a microscope; the total number of spores and number of sprouts were recorded, and the spore germination rate was calculated (Li et al., 2011).
Statistical analyses
All data were subjected to one-way analysis of variance to determine the significance of individual differences at the P < 0.05 level. Significant means were compared using the least significant difference (LSD) test. All statistical analyses were conducted using the SPSS commercial statistical package (SPSS, Version 17.0 for Windows, SPSS Inc., Chicago, USA).
Results
Antagonistic effect
A total of 34 strains with different morphologies were isolated from healthy bark by tissue separation and named B1-B34. Five strains (B9, B18, B26, B30, and B34) with different inhibitory effects on the pathogenic fungus were screened by confrontation culture. The antagonistic effect of B18 was the best, with an inhibition rate of 85.4% (Table 1).
Identification of an antagonistic bacterium
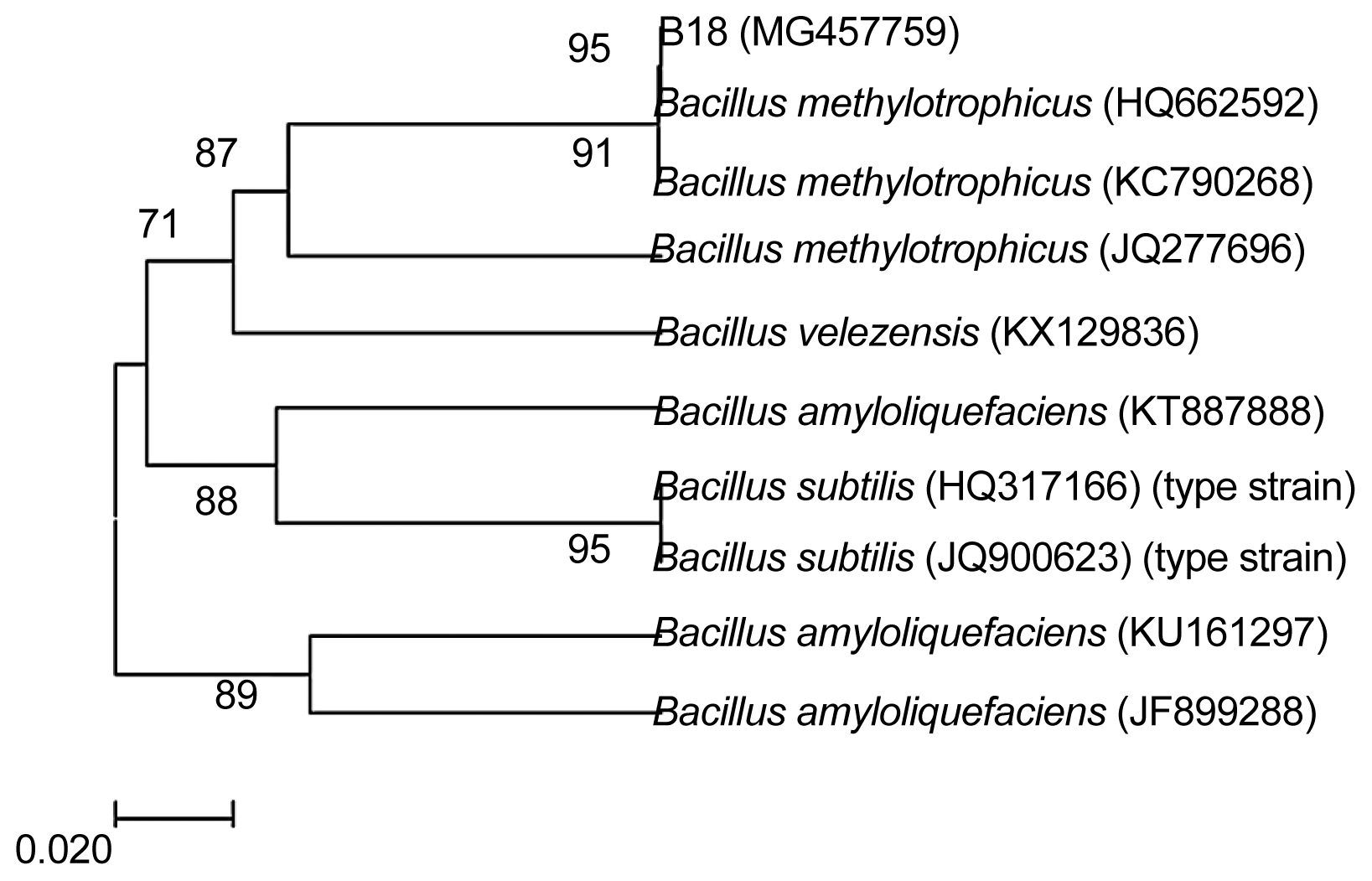
Morphological characteristics: Strain B18 was positive for Gram staining. The cell is a long rod in shape, with a size of 0.7-0.8 μm × 2.0-3.0 μm; endophytic spores formed are endophytic to secondary, elliptic or columnar, and slightly expanded (Supplementary Fig. 1). Physiological and biochemical properties: The results are shown in Table 2, referring to “Berger Manual of Bacterial Identification” (Fei et al., 2014). Strain B18 was identified as a member of the Bacillus genus and further identified by 16S rDNA sequence analysis. rDNA-ITS sequence analysis of antagonistic bacteria: 16S rDNA sequence primers (1492R/27F) of bacteria were used for PCR followed by agarose gel electrophoresis; the results are shown in Supplementary Fig. 2. The amplified product was sequenced, and an rDNA-ITS sequence of length 959 bp (GenBank accession number: MG457759) was obtained. The obtained sequence was analysed by homology BLAST with sequences reported in GenBank. A phylogenetic tree was constructed with MEGA5.0 analysis software (Fig. 1). The B18 strain and B. methylotrophicus (GenBank numbers: HQ662593 and KC790268) exhibit 99% nucleotide sequence similarity; a higher self-spreading value supports one branch far from other members of Bacillus. Therefore, strain B18 was identified as B. methylotrophicus.
Determination of control effects on potted plants
The control effects after inoculation with the B18 strain for 30 days in pot experiments are shown in Table 3, presenting symptoms such as those depicted in Supplementary Fig. 3. At a concentration of 104 cfu/ml, the cortex of P. amurense was nearly half rotten; leaves were dry, and a few were dead. Under the same conditions, bacterial inoculation in advance conferred the best control.
Field control
The control effects at 15 days after spraying treatment are shown in Table 4. The cure rate of the original was up to 96%. Better than 80% control was observed with concentrations of 108, 2 × 106, 5 × 105 and 2.5 × 105 cfu/ml, and the difference was not significant. The control effect of the concentration of 1.25 × 105 cfu/ml was significantly reduced to approximately 66%; while the disease was serious in the negative control group.
Antagonistic mechanism of the antagonistic strain
A growth curve of the pathogenic fungus (Fig. 2A) based on confrontation culture (Fig. 2B) was generated according to culture time, and the control group mycelium continued to increase in size. The colour of the mycelium and culture medium of the pathogenic fungus gradually deepened and darkened due to a control effect. Under an optical microscope, the morphology of the marginal mycelium of the inhibited colony was destroyed, mainly manifesting as swelling of the mycelium and deformity of the conidium. In contrast, the mycelia of the control group were vigorous and well developed (Fig. 2C). The results of the spore germination test are shown in Table 5. The B18 sterile culture filtrate demonstrated a distinct inhibitory effect on germination of the pathogen (Fig. 2D), whereas the spores in the control group were able to germinate normally. The germination rate was 77%.
Discussion
The beneficial bacteria of healthy plant tissue play an important role in controlling plant diseases. In this study, 34 strains of bacteria were isolated from the bark of healthy P. amurense, and 5 strains with different inhibitory effects against canker rot in P. amurense were screened by confrontation culture. The growth rate method was used to identify the best strain, B18, inhibiting the mycelial growth of canker rot in P. amurense. Morphological and molecular identification was carried out to examine the physiological and biochemical characteristics of this strain, and its control effects in potted plants and in the field were evaluated. The strain was identified as B. methylotrophicus by 16S rDNA analysis, and the strain showed strong antibacterial activity against the investigated pathogen. We also evaluated some interesting physiological and biochemical characteristics and biological activity. Bacillus-like bacteria can be isolated from the epidermis of healthy plant tissues, and many studies have shown that these microbes are antagonistic to many plant disease agents. However, biological control of B. methylotrophicus has not yet been studied. By investigating biocontrol effects, we can lay a theoretical foundation for the development of efficient and stable biological control agents.
The five strains exhibited more than 50% inhibition of the growth of N. haematococca. Strain B18, identified as B. methylotrophicus, inhibited the growth of N. haematococca mycelia. It has been reported that biocontrol bacteria can inhibit the growth of mycelia as well as the conidial germination of many plant pathogenic fungi in vitro, which is similar to the results of this study. A greenhouse test showed that inoculation with Bacillus grisea conferred 89.87% biocontrol, and the results of field experiments indicated that the filtrate of B. methylotrophicus had the highest fungistatic effect against Aspergillus oryzae, with 84.8% biological control (Shan et al., 2013). In this study, pot experiment showed better disease prevention with increasing concentrations of strain B18, with the best effect at 108 cfu/ml inoculation concentration. Similarly, in our field control analysis, the cure rate of canker rot in P. amurense was up to 96%, and the preventive effect decreased with increasing dilution of B18. In the pot and field trials in this study, B. methylotrophicus was inoculated in advance, and then the pathogenic fungus responsible for canker rot was inoculated; the best control effects were 98% and 96%, respectively, demonstrating broad prospects for development. Sun et al. (2016) found that B. methylotrophicus could control Microcystis aeruginosa and that the supernatant of the strain disrupted the cell membrane via oxidation, affecting membrane permeability; this bacterium has been used in an algaecide. Frikha-Gargouri et al. (2017) found that B. methylotrophicus had antimicrobial activity against Agrobacterium tumefaciens. In vitro, it was found that B. methylotrophicus could inhibit the growth of tomato crown gall disease pathogens.
Several studies have shown that B. methylotrophicus can produce a variety of metabolites with different functions. Zhang et al. (2014) found that B. methylotrophicus produces levosucrase and levopectin, and Chandankere et al. (2013) reported that B. methylotrophicus produces a biosurfactant with excessive foam formation. B. methylotrophicus strain USTBa represents an effective producer of biosurfactants and can be used in various biotechnology and industrial processes, especially in the petroleum industry. Chandankere et al. (2014) also used crude oil as the only carbon source and separated this effective bioactive agent-producing and hydrocarbon-degrading bacterium from the hydrocarbon pollution. Peng et al. (2015) was the first to prove that B. methylotrophicus can produce high levels of gamma-polyglutamic acid (γ-PGA), which has a great moisturizing effect and is not harmful to humans and accordingly is widely used in cosmetic manufacturing. Pérezflores et al. (2017) found that the increase in volatile production by B. methylotrophicus can lead to an increase in auxin content in Arabidopsis thaliana. In the present study, the growth rate method was used to determine the strong inhibitory effect of the B. methylotrophicus fermentation culture filtrate on the hyphal growth of N. haematococca mycelia. Strain B18 exhibited high biocontrol ability, meeting the screening criteria of an ideal biocontrol agent, and has a certain development potential. This study is a solid first step in developing an effective biocontrol agent for canker rot in P. amurense, but the exact mechanism by which the biological control bacterium strain B18 suppresses the growth of the pathogenic fungus remains to be further studied.

 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement
Supplement Print
Print






