

Fusarium head blight (FHB) is one of the most severe diseases threatening the production of major cereal crops including wheat, barley, and rice. Infected grains are often contaminated with mycotoxins such as zearalenone and trichothecences that cause mycotoxicosis in humans and animals (Desjardins, 2006; Lee et al., 2009). Although several fungal species of the Fusarium genus can cause FHB, the ascomycete fungus F. graminearum (telomorph: Gibberella zeae) is a major causal agent for FHB in Asia and North America (Leslie and Summerell, 2006). In Korea, F. graminearum recently caused severe yield losses in wheat, barley, rice, and maize (Lee et al., 2009, 2010, 2012; Ryu et al., 2011). In addition to yield losses and mycotoxin contamination through disease, planting infected seeds can cause root rot and seedling blight (Leslie and Summerell, 2006).
Despite the toxigenic and pathogenic importance of FHB, there are no FHB resistant cultivars and there is only limited information on biological agents against FHB. Fungicide application has shown partial efficacy for FHB control. Azole-type fungicides, one of the most important classes of systemic site-specific fungicides, reduce grain infection and trichothecene contamination (Pirgozliev et al., 2003). The fungicides inhibit cytochrome P450 sterol 14α-demethylase, resulting in the disturbance of fungal membrane integrity (Buchenauer, 1987). However, resistance to azole fungicides occurs in many plant pathogens including F. graminearum (Becher et al., 2010), limiting their widespread application and requiring the development of other approaches including resistant cultivars, cultivation methods, and biological control agents.
One of the most plentiful natural resources for the development of biological agents against plant pathogens are the Gram-positive Actinobacteria, which have high guanine and cytosine content in their genome. Stretomyces, which has highly differentiated branched mycelia, is the largest genus of Actinobacteria (Ventura et al., 2007). Many species belonging to this genus produce diverse secondary metabolites, some of which have antibiotic effects and affect soil microbial communities. In agriculture, the genus Streptomyces plays important roles in increasing soil fertility, degrading organic matter, and controlling pathogens. For example, this genus is a major soil decomposer that provides natural fertilizers for plants. Additionally, blasticidin S produced by S. griseochromogenes is effective for rice blast disease caused by the filamentous fungus Magnaporthe oryzea, and polyoxin produced by S. asoenosis is used for the control of bacterial rice sheath blight and fungal leaf spot (Watve et al., 2001). The diversity of Streptomyces species in soil provides numerous possibilities to explore potential biological control agents and new secondary metabolites that have antagonistic effects against plant pathogens.
Several filamentous fungi, yeast, and bacteria have antagonistic effects against F. graminearum (Luz et al., 2003; Pirgozliev et al., 2003) such as in vitro antifungal activity (Chan et al., 2003; Xue et al., 2009), the reduction of disease severity (Nourozian et al., 2006; Stockwell et al., 2001; Xue et al., 2009), systemic movement within infected spikes (Yuen et al., 2003), mycotoxin accumulation (Dawson et al., 2004), and pathogen survival (Bujold et al., 2001). Although many studies have focused on the development of biological agents against this disease, only a few biological agents including Bacillus subtilis and Cryptococcus laurentii strains have been evaluated under diverse growing conditions and developed as commercial biological agents (Pryor et al., 2007; Zhang et al., 2005). Therefore, more biological agents from natural environmental conditions need to be found and applied as alternatives to chemical approaches for control of FHB.
We found a bacterial colony that inhibited the mycelial growth of F. graminearum, suggesting that the bacterial strain can be developed as a biological agent against F. graminearum. In the present study, we identified the strain based on the sequences of the 16S rRNA gene and internal transcribed space (ITS) region as well as its morphological characteristics, and tested the efficacy of the strain against F. graminearum both in vitro and in vivo. Wheat heads treated with the BN1 strain spore suspension had significantly reduced FHB disease severity caused by F. graminearum, showing that the BN1 strain was able to control FHB.
Materials and Methods
Isolation and identification of the bacterial strain BN1
A bacterial colony was isolated from rice kernels that were placed on potato dextrose agar (PDA; Leslie and Summerell, 2006) to purify F. graminearum isolates. The bacterial colony was grown on lysogeny broth (LB; Sambrook and Russell, 2001) agar plate. To obtain chromosomal DNA, the strain, designated as BN1, was incubated in 3 ml of LB broth for 48 h at 28°C in an orbital shaker (200 rpm). Chromosomal DNA was isolated using the NucleoSpin columns (Macherey-Nagel, Duren, Germany) following manufacturer’s instruction. The primers ITS-38R (5′-CCG GGTTTCCCCATTCGG-3′) and ITS-72F (5′-TGCGGCTG GATCTCCTT-3′) (Normand et al., 1996) were used for amplification of the ITS region, and 16S-530F (5′-TGACT GACTGAGTGCCAGCMGCCGCGG-3′) and 16S-1494R (5′-TGACTGACTGAGGYTACCTTGTTACGACTT-3′) primers were used for amplification of the 16S rRNA gene (Borneman and Triplett, 1997). The amplified 16S rRNA gene was directly purified using MEGAquick-spinTM Total Fragment DNA Purification Kit (iNtRON Biotechnology, Sungnam, Korea) and directly sequenced. In the case of ITS region, the purified PCR product was cloned in pGEM-T Easy vector (Promega, Madison, WI), and the clone were sequenced. All the amplified DNA fragments and plasmid clones were sequenced using an automated 3730XL DNA sequencer (ABI, Tokyo, Japan). The sequences were used for a BlastN search in the NCBI GenBank database (http://blast.ncbi.nlm.nih.gov/Blast.cgi).
Phylogenetic analysis
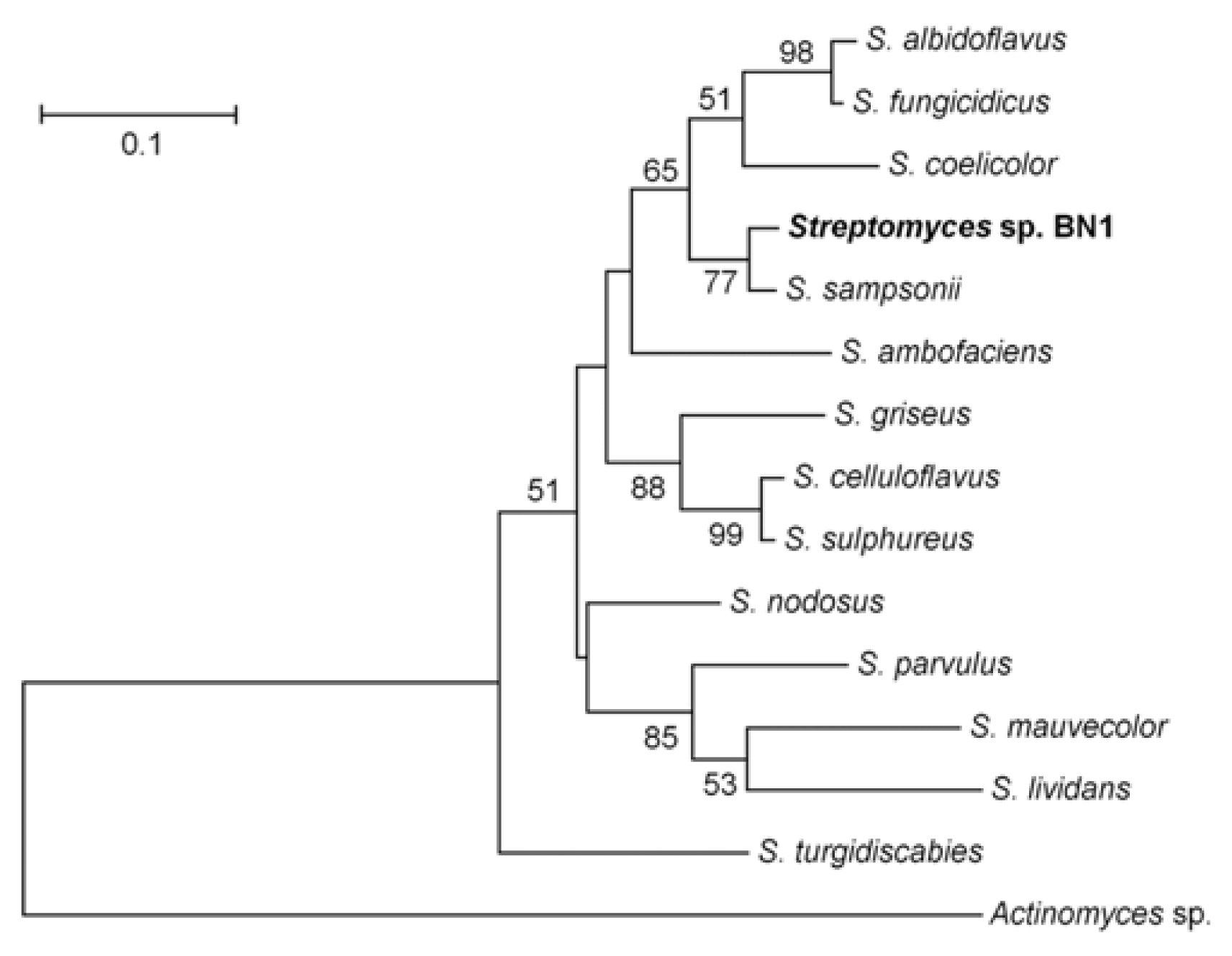
ITS sequences from the 13 Streptomyces spp. were retrieved from GenBank database. The sequences were aligned using the MEGA4.1 software with default parameters (Tamura et al., 2007), and trimmed manually at the same position that only includes ITS region. The ITS sequence of Actinomyces sp. strain F0330 (GenBank accession number NZ_ACTB01000202) was used as the outgroup for the phylogenetic anlaysis. A phylogenetic tree was constructed using the neighbor-joining method (Saitou and Nei, 1987) in the MEGA4.1 software with the following parameters: complete deletion of gaps, Kimura-2, both transitions and transversions substitutions included, and 2,000 bootstrap replicates.
Preparation of inoculum
The bacterial strain BN1 was incubated in 30 ml of LB for 72 h at 28°C in an orbital shaker (200 rpm) to induce spore production. To harvest BN1 spores, the culture was filtered with two layers of miracloth (Calbiochem, La Jolla, CA, USA) and the filtrates were centrifuged at 10,000 g. The spores were washed twice with distilled water and resuspended with 0.1% (v/v) Tween 20 solution (106 spores/ml). To induce spore production of F. graminearum strain JL001, the strain was incubated in 30 ml of carboxymethylcellulose (CMC) medium as previously described (Capellini and Peterson, 1965). The cultures were also filtered with two layers of miracloth and spores were harvested by centrifugation at 10,000 g. Spores were washed twice with distilled water and diluted to a final 105/ml concentration in 0.1% (v/v) Tween 20 solution.
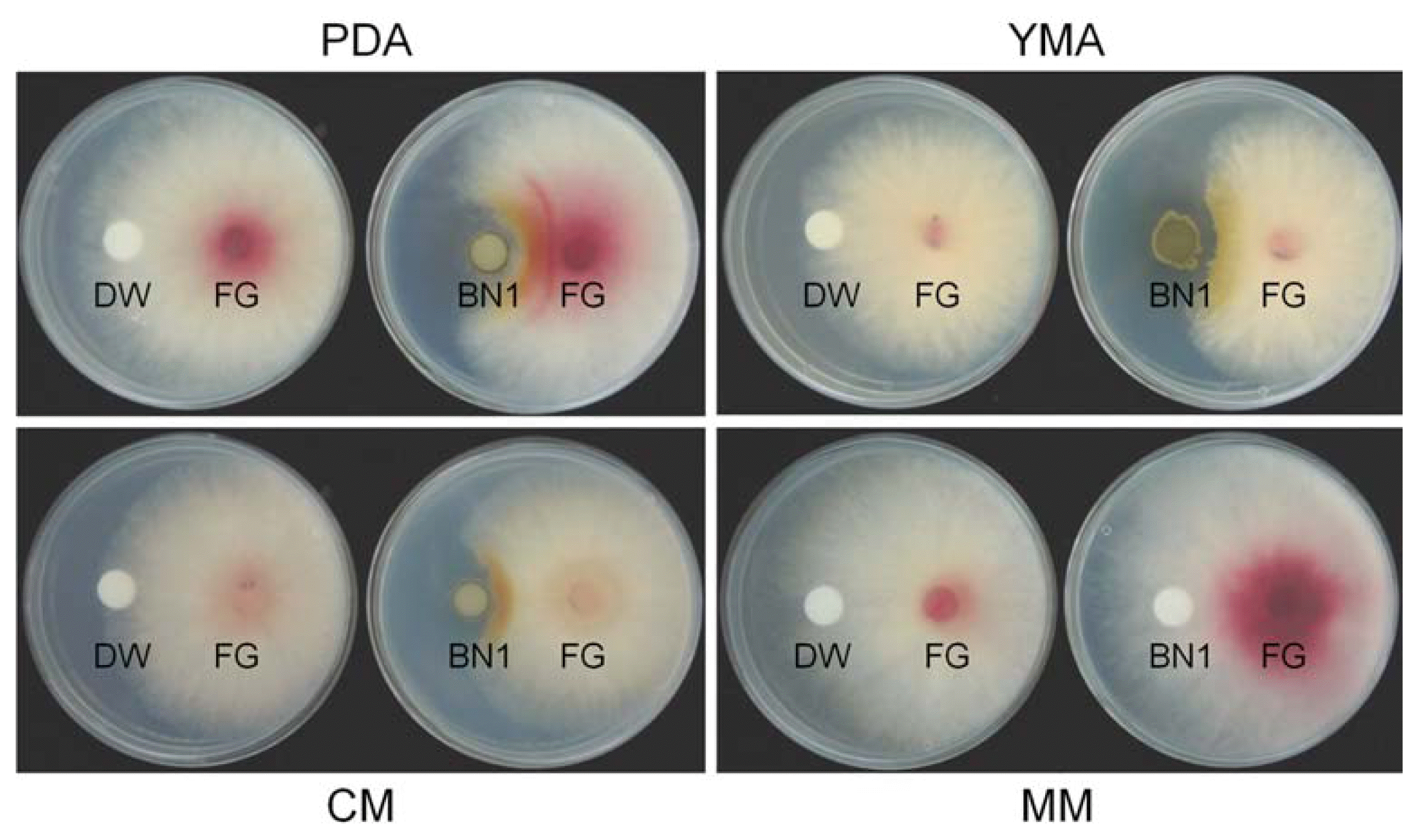
Antifungal activity assay of the bacterial strain BN1 on diverse media
The biological efficacy of the BN1 strain against F. graminearum was tested on four different media including PDA, yeast malt agar (YMA; Harris, 2005), complete medium (CM), and minimal medium (MM; Leslie and Summerell, 2006). A sterilized paper disk was placed on one side of each 9-cm Petri-dish and 20 μl of BN1 spore suspension (106 spores/ml) or distilled water dropped onto the paper disk. An agar block (5 mm diameter) taken from the margin of a 5-day-old PDA culture of F. graminearum JL001 strain was placed on the other side of each plate. The inoculated plates were incubated at 25°C for five days. To check the inhibitory effect of the bacterial strain against the fungal strain, distance between the bacterial colony and fungal colony was measured.
Efficacy assays of BN1 against F. graminearum on wheat seedlings
Wheat cultivar Eunpamil, which is susceptible to FHB, seeds were soaked in distilled water or BN1 spore suspension (106 spores/ml) for 1 h in an orbital shaker (100 rpm). The wheat seeds were dried on a clean hood for 2 h and then each ten seeds were planted in pots. Before covering the seeds with soil, one ml of F. graminearum conidia suspension (105 spores/ml) was sprayed onto the seeds. The pots were placed in a greenhouse for 2 weeks and the length of each seedling was measured to determine the efficacy of BN1 against F. graminearum during wheat seedling growth. All experiments were repeated three times and the Tukey test using SPSS 12.0 software (SPSS Inc., Chicago, IL, USA) was performed to determine the significant differences (P < 0.05) among the mean values of samples.
Efficacy assays of BN1 against F. graminearum on wheat heads
The efficacy of BN1 against F. graminearum on wheat heads was tested using two inoculation methods including spraying and point inoculation. For the spraying inoculation, one ml of F. graminearum spore suspension (105 spores/ml) or one ml of mixed spore suspension (105 of F. graminearum spores and 106 of BN1 spores) were sprayed onto each head (at the mid-anthesis stage) of the wheat cv. Eunpamil. For the point inoculation, 10 μl of F. graminearum spore suspension (105 spores/ml) or 10 μl of mixed spore suspension (2 × 105 of F. graminearum spores/ml and 2 × 106 of BN1 spores/ml) were injected into a center spikelet of a wheat head at mid-anthesis. For the negative controls, wheat heads were treated with sterilized distilled water only or BN1 spore suspension without F. graminearum spore suspension. The inoculated wheat head was wrapped with a plastic bag and maintained for 2 days, after which the plastic bags were removed from the infected wheat heads. The plants were placed and grown in a greenhouse and the number of wheat kernels was counted after 2 weeks.
Results
Identification of the bacterial strain BN1
During survey of rice kernels infected by FHB, we isolated a bacterial colony, named BN1. The bacterial strain BN1 produced branched hyphae and spores both on solid and in liquid LB, CM, and YMA media. Aerial hyphae on solid media often had long, straight filaments bearing spores. These morphological characteristics suggest that this strain belongs to the genus Streptomyces. The 16S rRNA primers amplified a 956 bp fragment from genomic DNA of the BN1 strain and the amplified fragment had 100% sequence identity with the 16S rRNA gene found in several Streptomyces spp. including S. albidoflavus, S. gougerotii, S. rutgersensis, S. diastaticus, and S. rutgersensis. This result confirmed that the BN1 strain belonged to the genus Streptomyces, but the 16S rRNA was not diverse enough to identify the species level in this genus. ITS-38R and ITS-72F primers amplified a 474 bp fragment. We excluded the 16S and 23S rRNA regions from the amplified sequence and finally used a 321 bp ITS region to compare the ITS sequences to other known bacterial species. The ITS region of the BN1 strain showed the highest identity (96%) with that of S. sampsonii, and the phylogenetic tree showed that BN1 grouped with S. sampsonii (77% BS) (Fig. 1). The alignment and trimming of ITS region sequences produced 331 bp with gaps, of which 177 sites (53.5%) were conserved and 150 sites (45.3%) were variable. The overall mean distance of the applied sequences was 0.246. In the NJ tree, all the 13 Streptomyces spp. and Streptomyces sp. BN1 showed distinct differences (Fig. 1). Eleven sites (3.4%) were variable among the 320 sites of ITS region sequences between between BN1 and S. sampsonii (data not shown).
In vitro efficacy of the BN1 strain against F. graminearum.
The in vitro efficacy level of BN1 strain against F. graminearum was slightly different based on the media tested. The BN1 strain mostly inhibited F. graminearum mycelia growth on YMA, PDA, and CM, but not on MM (Fig. 2). The level of inhibition in the media was dependent on the extent of BN1 colonization on each medium. On MM, BN1 strain did not grow at all, resulting in the non-efficacy of BN1 against F. graminearum on this medium (Fig. 2). When the BN1 strain was first inoculated in PDA, YMA, and CM to allow colonization in the media, followed by the inoculation of F. graminearum 3 days later, the inhibition levels were higher than co-inoculation of BN1 and F. graminearum (data not shown).
Efficacy of BN1 on wheat seed germination and seedling growth
Wheat seeds soaked in or sprayed with F. graminearum spore suspension had a similar germination rate as wheat seeds that were not treated with F. graminearum spore suspension. All experimental conditions had 70-80% wheat seed germination rate. However, seedling length was significantly decreased in the plants inoculated with F. graminearum spore suspension compared to that of non-inoculated plants (Table 1). Seedling length of the wheat seeds treated with BN1 spore suspension before F. graminearum inoculation was not significantly different from the non-inoculated plants (Table 1).
Efficacy of BN1 against F. graminearum on wheat heads
When the spore suspension was sprayed onto wheat heads, the F. graminearum strain JL001 produced typical FHB symptoms after a 14-day inoculation, while BN1 did not trigger any disease symptoms (Fig. 3 and Table 1). When the F. graminearum spore suspension was sprayed onto wheat heads with the BN1 spore suspension, the disease severity of FHB was significantly reduced compared to that of FHB caused by only F. graminearum inoculation. However, when the wheat heads were inoculated by the point-inoculation method, BN1 did not have any efficacy against F. graminearum (Fig. 3 and Table 1).
Discussion
Epidemics of FHB have re-emerged worldwide (Xu and Nicholson, 2009), and frequent rainfall during the wheat flowering season in 2011 in Korea resulted in FHB epidemics and fatal yield losses in wheat production. Even after reducing the light wheat kernels that might be infected by F. graminearum, germination rate of wheat seeds was below 70%, resulting in a shortage of seeds for planting that year. In spite of the drastic yield losses in wheat production, there are neither resistant cultivars against this disease nor registered fungicides for seed treatment and field application in Korea. These severe situations require the development of biological agents against FHB, and the present study provides one potential biological agent purified from a rice field.
Recently, the population structure of F. graminearum field strains in Korea have been extensively studied based on molecular markers and chemical analyses (Lee et al., 2009; 2010; 2012). The Fusarium species complex consists of at least 14 phylogenetic lineages based on the genealogical concordance (O’Donnell et al., 2000), and recently each lineage was classified as a new species (O’Donnell et al., 2004; 2008). The geographic location often influences the lineage distribution. For example, lineage 7 (F. graminearum sensu stricto) is predominant in North America and lineage 6 (F. asiaticum) is predominant in Asian regions including Korea, China, and Japan (Gale et al., 2002; Lee et al., 2009; 2012; Suga et al., 2008). In Korea, the population structure is more complicated than that of other countries. In southern provinces of Korea, lineage 6 is predominant in rice and wheat. In the middle-eastern provinces, lineage 6 is predominant in rice fields, but lineage 7 is predominant in maize fields. Lineage 6 may be predominant in rice fields of Korea because it is better suited to a rice agro-ecosystem than the other lineages present in Korea (Lee et al., 2009). In addition, F. graminearum strains in Korean rice and maize fields have different genetic characteristics compared to those of other countries (Lee et al., 2009, 2012). The populations have lower genetic and genotypic diversity, suggesting that the populations have been maintained through asexual and homothallic events. These studies suggest the need to develop approaches to maintain the Korean F. graminearum populations.
Korea predominantly uses organic wheat production, and no chemical fungicides or biological agents have been registered for controlling wheat diseases or seed treatment. Therefore, controlling diseases in Korean wheat fields has focused on cultivation techniques, which have not been sufficient to control FHB epidemics. Frequent epidemics of FHB in Korea have led to the registration of fungicides such as prothioconazole, tebuconazole, and metconazole that have been widely used worldwide (Pirgozliev et al., 2003). However, frequent usage of the fungicides has triggered the occurrence of F. graminearum strains resistant to the fungicides (Becher et al., 2010). The registration and application of fungicides in Korea could possibly control FHB in Korea for a few seasons, but fungicide-resistant strains will likely develop. Therefore, we need to develop alternative methods of control, including finding biological agents against F. graminearum.
Microorganisms have been used for development of biological agents against various plant pathogens, particularly the bacterial genus Streptomyces that has more than 600 species and produces various kinds of secondary metabolites with antagonistic effects to other microbes (http://www.bacterio.cict.fr/s/streptomycesa.html). In this study, a new Streptomyces species was isolated from a Korean rice field and the strain showed a highly antagonistic effect against F. graminearum both in vitro and in vivo. The sequences of 16S rRNA genes in the genus Streptomyces are highly conserved and are only useful for classification at the genus but not species level. Sequencing analysis showed that the 16S rRNA gene sequence of the BN1 strain matched morphological characteristics belonging to the genus Streptomyces. The ITS sequence of BN1 strain was different from those of known Streptomyces species. The phylogenetic tree showed that the ITS sequence of BN1 grouped with that of S. sampsonii with 77% BS, suggesting that BN1 does not belong to any known Streptomyces species. To classify the BN1 strain, further studies in morphological characteristics, phylogenetic analyses, and serological studies are needed.
The initial infection of F. graminearum occurs by ascospores discharged from perithecia during the short period (10-20 days) from anthesis through the soft dough stage of kernel development (Schroeder and Christensen, 1963). Infections are most serious when the anthers are exposed to 25-30°C with high humidity during flowering (Parry et al., 1995; Sutton, 1982). To verify the efficacy of the BN1 strain against F. graminearum, we directly inoculated wheat heads with spore suspension during mid-anthesis and maintained humidity for 2 days. Both spraying and point inoculation successfully caused FHB symptoms in wheat heads. However, the BN1 strain only efficiently antagonized F. graminearum with the spraying method, possibly because of either colonization of the BN1 strain within anthers or movement of the BN1 strain between anthers. Spores of F. graminearum germinate within anthers and move to another anther to cause blight symptoms throughout wheat heads. The BN1 strain did not trigger any disease symptom in wheat heads, suggesting that it is saprophytic on wheat plants, and the colonization of BN1 may be later than that of F. graminearum within a limited area such as wheat anthers. Also, the mycelial growth of F. graminearum is faster than that of BN1, and F. gramineruam efficiently overcomes the efficacy of BN1. Importantly, BN1 had an antagonistic effect using the spraying inoculation method, which is more natural than the point inoculation method for FHB development in wheat heads. This result suggests that the BN1 strain grew better on the wheat heads than in anthers, and that wheat heads can be successfully protected from F. graminearum infection by spraying BN1 spores during flowering.
In this study, a Streptomyces species strain BN1 was isolated from a rice field in Korea. The BN1 strain was identified as a new species based on the phylogenetic analysis of the ITS sequence. The efficacy of the BN1 strain against F. graminearum was characterized both in vitro and in vivo. BN1 efficiently inhibited the mycelial growth of F. gramienarum on media, and co-inoculation of F. graminearum and BN1 spore suspension into wheat heads reduced FHB disease severity. This study provides one of the best potential strategies for FHB control in Korea that could guarantee sustainable wheat production in Korea. Further studies are needed on the classification of BN1 at the species level and the mechanism of BN1 inhibition against F. graminearum.

 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print






