



Heilongjiang province, situated in the northeast of China, is recognized as the birthplace of soybean cultivation and a major contributor to the global soybean production (Hymowitz and Shurtleff, 2005). The region’s black soil, with its dark color and crumbly texture, is renowned for its fertility and nutrient richness, accumulated over thousands of years of organic matter (Zhao et al., 2014). Moreover, the high water-holding capacity of Heilongjiang’s black soil makes it an ideal place for cultivating different crops, including soybeans, corn, wheat, and rice (Cai et al., 2022b). Additionally, the region’s soil harbors a diverse range of microorganisms that play a crucial role in its ecosystem (Zhou et al., 2017).
Despite being rich in nutrients and microorganisms, the soil in Heilongjiang has been undergoing severe degradation over the years, primarily due to monoculture planting systems and the extensive use of fertilizers and pesticides (Du et al., 2021, Mei et al., 2022). This has caused soil compaction and a decline in biodiversity, allowing certain soil-borne pathogens like Fusarium spp. to dominate (Cai et al., 2022a; Husaini et al., 2018). The prevalence of Fusarium presents a new threat to soybean production, making it imperative to conduct a comprehensive investigation of its distribution in the region and develop better management strategies to combat the pathogen.
The genus Fusarium is notorious for housing numerous plant pathogenic fungi, and several species within this genus have been known to cause severe soybean diseases globally, such as root rot, sudden death syndrome, blight, wilt, and damping off (Arias et al., 2013; Cai et al., 2020; Lin et al., 2022). While using fungicidal seed treatment is a common approach to managing Fusarium infection in soybean, it is not environmentally friendly and can lead to soil degradation and reduced soil biodiversity (Ayesha et al., 2021). An alternative solution is to cultivate soybean varieties that are resistant to Fusarium, but this requires a thorough understanding of the distribution and virulence of the different Fusarium species.
To improve the management of Fusarium diseases in soybean in Heilongjiang province, a two-year survey of soybean root rot was conducted. The study aimed to identify the Fusarium species responsible for soybean root rot, assess the virulence of these species on soybean, and evaluate the resistance of soybean cultivars to the major Fusarium species. The information gathered from this study will facilitate the development of more effective management strategies against Fusarium in the region and promote sustainable soybean production in Heilongjiang province.
Materials and Methods
Sample collection
For agricultural production purposes, Heilongjiang Province has been classified into six accumulated temperature zones (ATZs), with each zone utilizing specific soybean cultivars to achieve optimal yield (Lu et al., 2017). Over a period of two years, soybean roots affected by the disease were sampled, resulting in the collection of 206 root samples from across the six ATZs, which represent the primary soybean production areas.
Isolation of Fusarium species
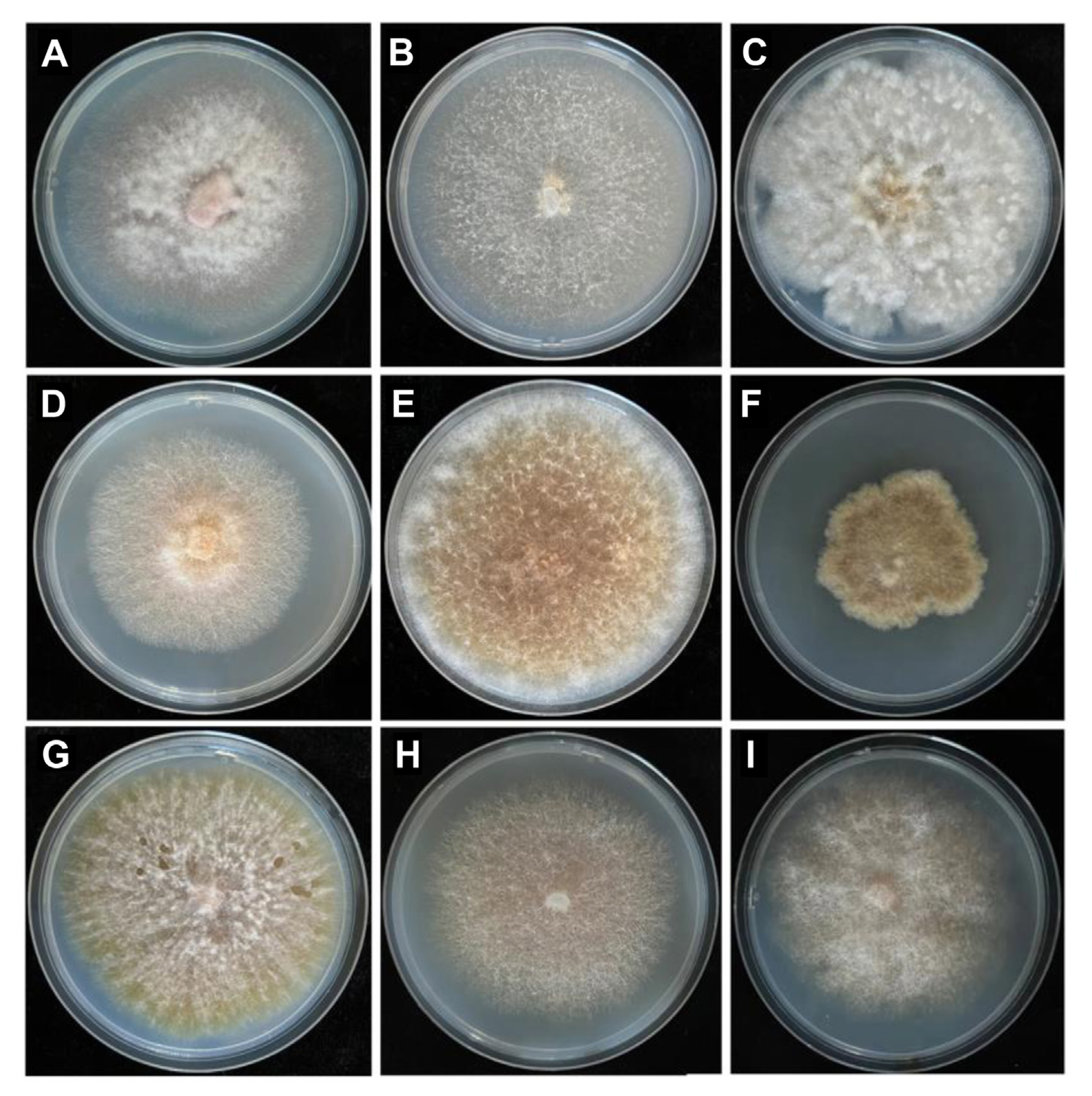
The pathogen was isolated using tissue separation method. First, the diseased plant samples were washed with sterilized distilled water. Then, the infected root tissues of the plants were cut into small pieces of about 1 cm, and sterilized with 75% ethanol for 1 min. After rinsing with sterile water for 5 times, they were treated with 10% sodium hypochlorite for 5 min, and then rinsed with sterile water for 5-8 times until clean. The surface moisture of the tissue was absorbed with sterilized filter paper. Root pieces were embedded in antibiotic-amended Nash-Snyder medium (15 g peptone, 1 g KH2PO4, 0.5 g MgSO4·7H2O, 20 g agar, 0.75 g PCNB, 33 mg/ml streptomycin sulfate solution, 40 mg/ml neomycin sulfate solution per liter) and incubated at a constant temperature of 28°C for 5 days. After the fungal colonies grew, the isolates were purified by single-spore isolation and grown on potato dextrose agar for morphological identification, molecular identification, and pathogenicity testing.
DNA extraction, polymerase chain reaction amplification, and sequencing
DNA extraction and purification were performed as described previously (Arias et al., 2013). Briefly, the mycelium of each isolate was collected by scraping the surface of growing colonies on potato dextrose agar medium incubated for 1 week at 28°C. The fungal mycelia were ground in liquid nitrogen, then the genomic DNA was extracted using the Fungal DNA mini kit (Tiangen Biotech, Beijing, China) as described by the manufacturer. The DNA purity and concentration were determined by using a NanoDrop ND1000 spectrophotometer (NanoDrop Technologies Inc., Waltham, MA, USA). The complete rDNA-ITS and the highly conserved mitochondrial and nuclear ribosomal small subunit (mtSSU rRNA genes) of Fusarium were amplified. The primers used were ITS1/ITS4 (5′-TCCGTAGGTGAACCTGCGG-3′, 5′-TCCTCCGCTTA TTGATATGC-3′) and MS1/MS2 (5′-CCAGCAGTCAAGAATATTAGTCAATG-3′, 5′-GCGGATTATCGAATTAAATAAC-3′), respectively (O’Donnell et al., 1998; White et al., 1990). These primers were synthesized by Sangon Biotech (Shanghai) Co., Ltd. Polymerase chain reaction (PCR) products were sequenced at the Sangon Biotech (Shanghai) Co., Ltd. Sequences were submitted for BLAST searches for comparison to known DNA sequences in the NCBI. Random amplified polymorphic DNA (RAPD)-PCR was carried out in a total volume of 25 μl containing 12.5 μl 2× SanTaq PCR mix, 8.5 μl double distilled water, 1 μl primer (10 pmol/μl) and 1 μl template DNA (40 ng/μl). Amplification was run with the following profile: an initial denaturation at 94°C for 5 min, followed by 45 cycles of denaturation at 94°C for 30 s, annealing at 36°C for 1 min, extension at 72°C for 2 min, and a final extension at 72°C for 10 min.
Pathogenicity of Fusarium species
For pathogenicity assay of nine Fusarium species, soybean cv. Xiaojinhuang was used. Soybean seeds were sterilized with 75% ethanol for 3 min and then washed three times with sterile water. Two seeds were planted per pot containing a 3:1 mixture of sterile soil and vermiculite. Inoculation was performed about two weeks after emergence of plants, as described previously (Cai et al., 2020). Six strains from different ATZs were used for pathogenicity test for each Fusarium species. Spore suspensions for inoculation were prepared by flooding cultures with distilled water, then filtering them through sterile cloth. The final conidia concentration was adjusted to 106 conidia/ml. Nine plants per treatment were immersed in a spore suspension for 2 h. Then the roots were washed with distilled water and put back into the pots for further growth. Disease index was calculated according to Chang et al. with some modifications (Chang et al., 2015). Briefly, after 21 days of inoculation, soybean seedlings were gently removed from the pot by turning it upside down, the soybean plants are repeatedly rinsed with clean water, and then the surface of the plant is dried with filter paper. Based on the symptoms of the main and lateral roots and the degree of rot, soybean root rot caused by Fusarium is divided into five levels; Level 0: no disease symptoms; Level 1: the main root is lightly browned, the lateral root growth point is diseased, but the plant can grow normally; Level 2: the main root is diseased and turns from brown to black, the tip of the lateral root turns black, but the plant can still grow normally; Level 3: most of the main root turns black and cannot continue to grow, the lateral roots are reduced or there are no lateral roots, and the plant grows slowly; Level 4: the roots are completely rotten, and the plant cannot continue to grow and may even wilt and die. By observing the root disease situation of Fusarium interacting with soybean plants, referring to the grading standards, the disease grades of different Fusarium infections are recorded, and the disease index is calculated according to the formula as follows.
Disease index = ∑ (number of plants × number of degree in symptoms)/(total number of plants × 5(maximum degree in symptoms)×100. Plant height and root length was also measured, and the shoot dry weights and root dry weights recorded for each plant after oven drying for 24 h at 60°C. Three replications were conducted to assess the pathogenicity of each Fusarium isolate.
For the soybean cultivar resistance test, a total of 21 cultivars were included in the study. The specific cultivars used are HN48, HN51, HN53, HN64, HF35, HF47, HF51, DN44, DN51, DN54, DN57, HN63, HN75, BD48, SN30, SN31, SN35, SN37, LD5, KF21, and HL44. As the most frequent species identified in this study, F. oxysporum and F. solani were selected. Soybean seeds were sterilized with 75% ethanol for 3 min and then washed three times with sterile water. Inoculation was conducted by dipping 200 ml 106 conidia/ml spores to soybean seed surface. Then the seeds were put in dark condition for seven days. Disease severity was measured by dividing length of infected brown root to whole root length.
Results
Diversity of Fusarium species associated with soybean root rot
Over the course of a two-year survey, a total of 485 isolates belonging to the Fusarium genera were recovered, and nine species were identified based on characteristics of colony and spore structure, as well as ITS and mtSSU rRNA gene sequences (Fig. 1, Supplementary Figs. 1 and 2). Among all pathogenic Fusarium, a total of 227 strains of F. oxysporum were isolated, with an isolation frequency of 46.80%, ranking first in proportion, followed by F. solani with an isolation frequency of 19.38%. F. equiseti was the third most abundant species, with a relative abundance of 11.75%. The other Fusarium species identified were F. proliferatum, F. graminearum, F. tricinctum, F. brachygibbosum, F. globosum, and F. incarnatum. Among these, F. globosum was first discovered in China from diseased soybean plants with root rot. These findings highlight the diverse nature of Fusarium species present in the surveyed region.
Distribution of Fusarium species
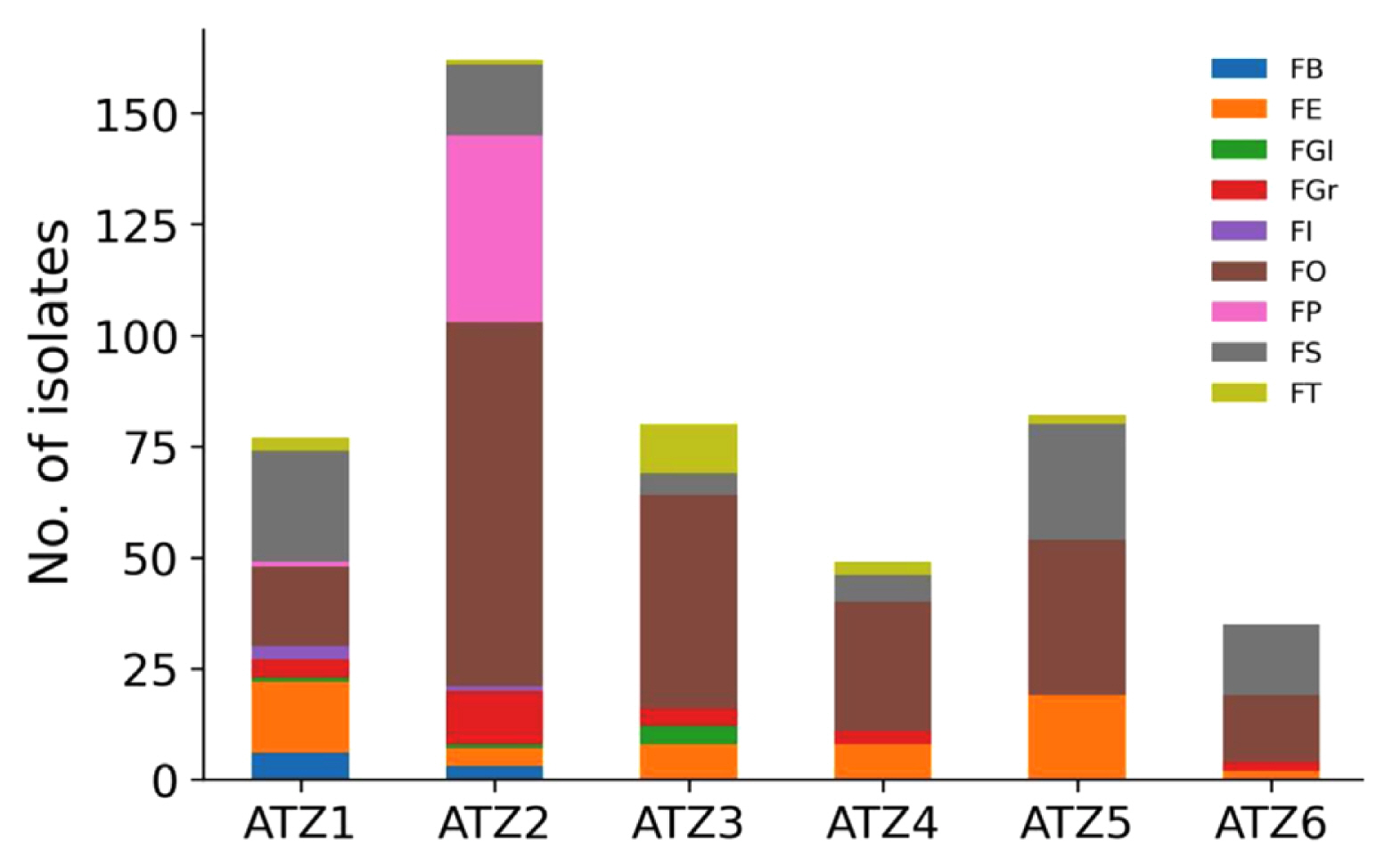
The distribution of the isolated strains in different ATZ is shown in Fig. 2. F. oxysporum, F. solani, and F. equiseti are distributed in all regions; F. brachygibbosum, F. incarnatum, and F. proliferatum are only distributed in the first and second ATZs; F. globosum is found in the first, second, and third ATZs; among the six ATZs, only F. tricinctum was not found in the sixth ATZ, while it is distributed in all other regions. F. graminearum was not found in the fifth ATZ among the six ATZs. The data also shows that the distribution of Fusarium species is significantly reduced in the fifth and sixth ATZs, the second ATZ has the most diverse distribution of Fusarium species and the highest number of pathogenic strains. F. oxysporum and F. solani were found to be the dominant species in almost all six ATZs, indicating their widespread distribution throughout the region. This suggests that these two species have the potential to cause significant damage to agricultural production and warrant further study. Intriguingly, ATZ2 had the highest abundance of Fusarium spp. in terms of both species diversity and quantity, suggesting that temperature plays a crucial role in the pathogen’s distribution. The result is in agreement with prior investigations that have illustrated the impact of temperature on the growth and survival of Fusarium spp. Understanding the distribution and environmental factors influencing the prevalence of Fusarium species is critical for developing effective strategies to control their spread and minimize the impact on crop yields.
RAPD analysis of 33 F. oxysporum isolates from six ATZs
Out of the 30 primers that were tested, only 10 RAPD primers yielded PCR products with clear and vibrant polymorphic bands. Conversely, the remaining 20 primers resulted in negligible band formation in the PCR products. The 10 different primers consistently amplified multiple polymorphic bands, ranging from 5 to 11 bands (Supplementary Fig. 3). In total, 79 RAPD markers were acquired, out of which 43 were polymorphic, constituting 54.43% of the total markers obtained (Table 1).
Pathogenicity assay of Fusarium species on soybean plant
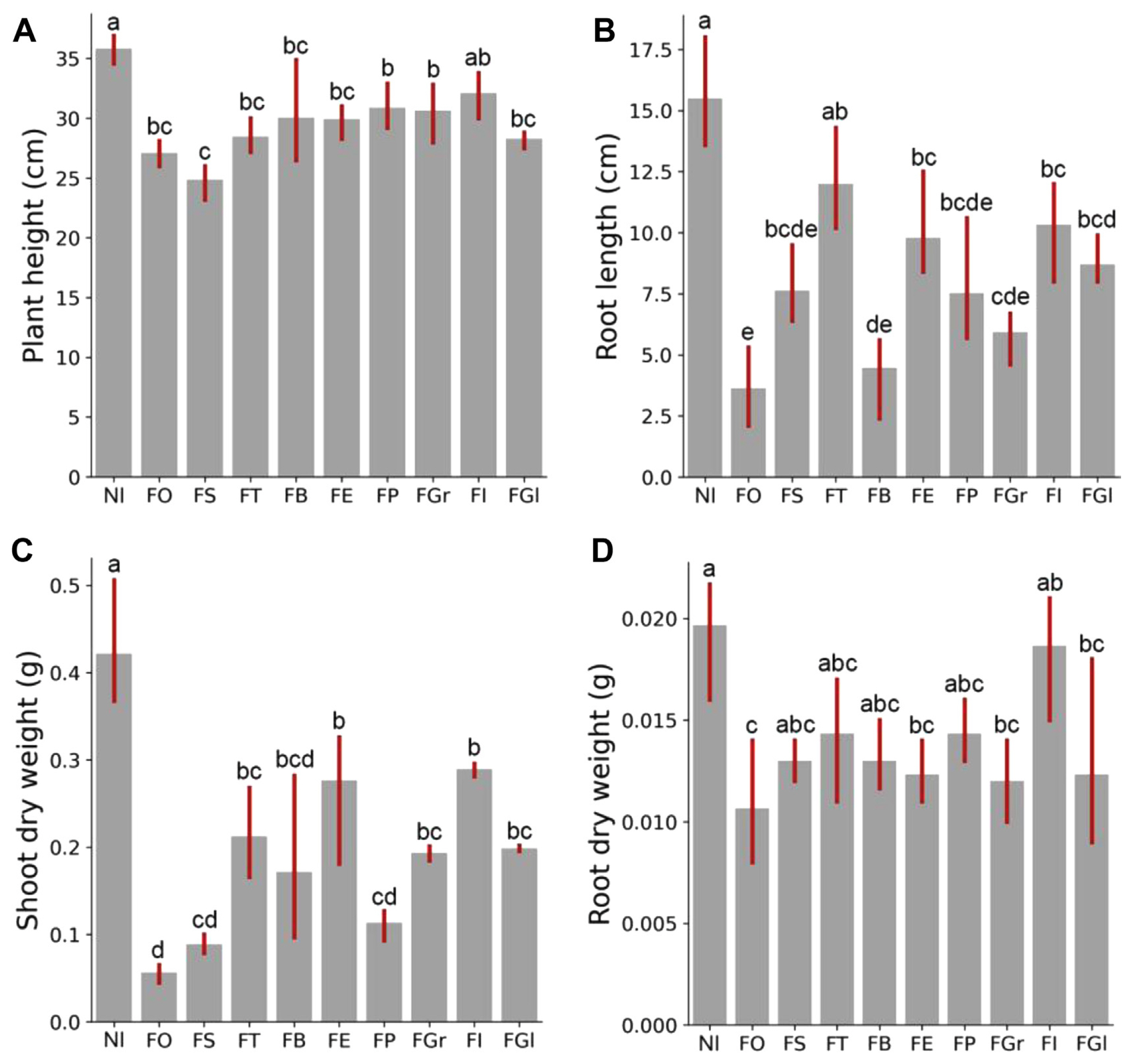
To examine the relative aggressiveness of different Fusarium species on soybean roots, a series of pot experiments were conducted. Nine Fusarium species were tested, and the results revealed significant differences in their ability to cause root rot (Fig. 3). F. oxysporum and F. solani were found to be the most aggressive pathogens, causing the most severe root rot symptoms and being the most frequently isolated species. F. tricinctum, F. proliferatum, F. graminearum, F. brachygibbosum, F. equiseti, F. globosum, and F. incarnatum also caused significant damage to plant growth and development, as evidenced by reductions in plant height, root length, shoot dry weight, and root dry weight compared to the control (Fig. 4). Further analysis of the data showed that the different Fusarium species had varying effects on plant characteristics. F. oxysporum was found to be the most severe species in terms of its impact on plant growth, indicating that it is the most aggressive isolate. These findings are consistent with previous studies that have shown F. oxysporum and F. solani to be highly virulent pathogens that can cause significant yield losses in soybean (Hafez et al., 2021).
Soybean cultivars responses to F. oxysporum and F. solani infection
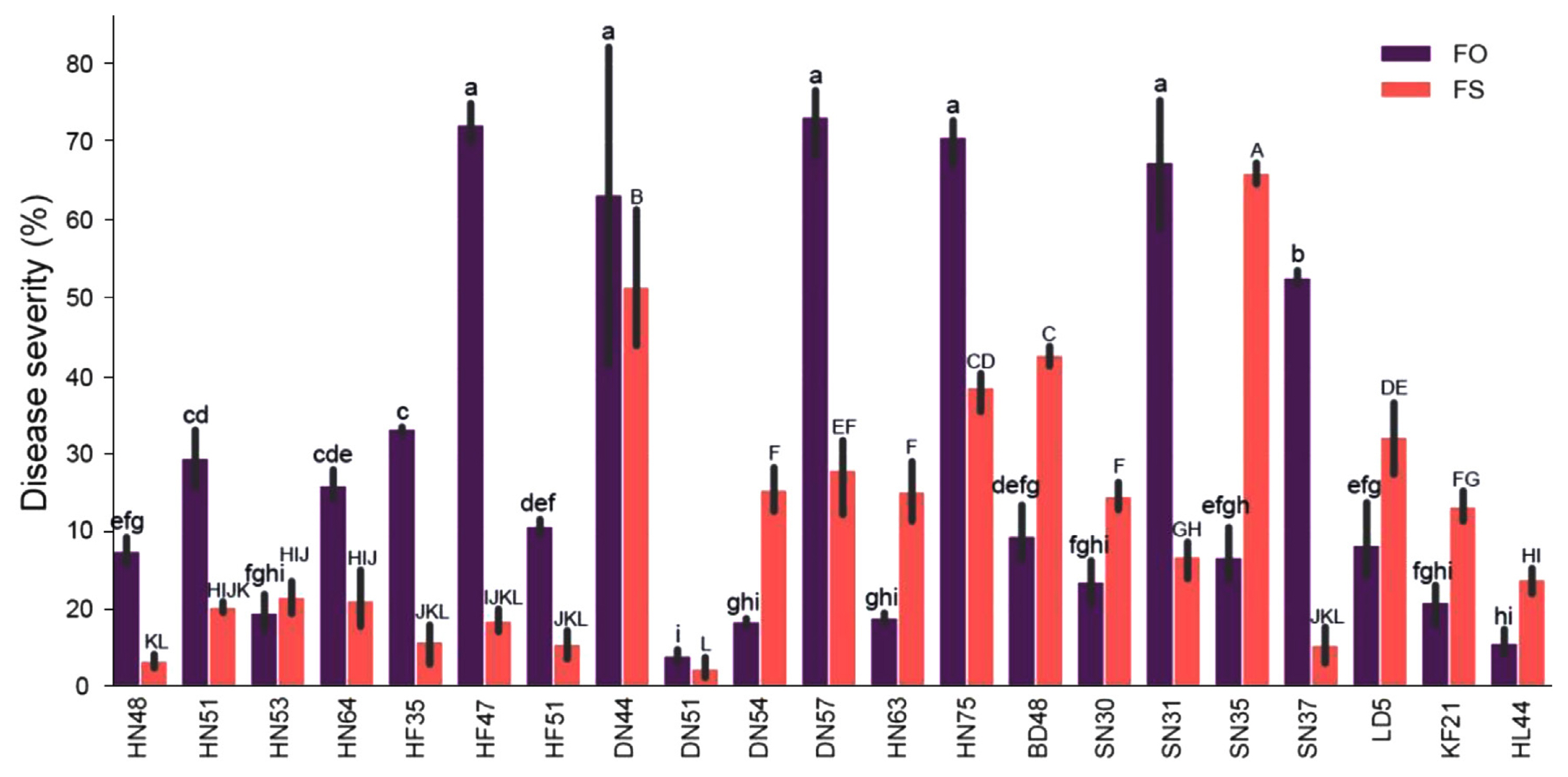
The results of disease susceptibility testing indicated significant variation in the response of different soybean cultivars to F. oxysporum and F. solani infections (Fig. 5). Most cultivars showed greater susceptibility to F. oxysporum infection than to F. solani infection, with F. oxysporum causing more severe disease symptoms. However, for some cultivars, including BD48, SN30, SN35, LD5, KF21, and HL44, F. solani infection resulted in more severe disease symptoms than F. oxysporum infection. Interestingly, there were no significant differences in disease severity between F. oxysporum and F. solani infection for cultivars HN53, DN44, and DN51. Moreover, the soybean cultivar DN51 showed the most resistance to both F. oxysporum and F. solani infection, indicating that it may possess genetic traits that make it less susceptible to Fusarium root rot.
Discussion
Pathogens and plants are not independent entities. Disease development requires the presence of three key factors: a susceptible plant host, a virulent pathogen, and favorable environmental conditions. If any of these factors are absent or unfavorable, disease will not occur (Velásquez et al., 2018). Numerous studies have shown that temperature can affect both pathogen growth and reproduction, as well as the severity of diseases caused by Fusarium spp. (Abbas et al., 2022; Cruz et al., 2019; Kaur et al., 2022). Climate has also been identified as a significant factor in determining the distribution of Fusarium spp. in soil worldwide, implying that temperature may play a critical role in the incidence and severity of Fusarium-caused plant diseases (Nikitin et al., 2023; Poole et al., 2013).
Climate parameters such as accumulated temperature during soybean growth, relative humidity, precipitation, sunlight duration, and soil surface temperature significantly impact the distribution of Fusarium pathogens. The occurrence of Fusarium root rot primarily occurs in warm and humid regions, with the severity of the Fusarium infection increasing with higher precipitation and relative humidity (Abbas et al., 2022). The climate in Heilongjiang province spans a large range, featuring limited heat resources and significant differences in accumulated temperature (Li et al., 2022). The first ATZ has an accumulated temperature of over 2,700°C and a warm climate. The second ATZ has a moderate climate with an accumulated temperature of between 2,500-2,700°C, the third ATZ has a cool climate with an accumulated temperature of between 2,300-2,500°C, the fourth ATZ has a cold climate with an accumulated temperature of between 2,100-2,300°C, the fifth ATZ has a very cold climate with an accumulated temperature of between 1,900-2,100°C, and the sixth ATZ has a high and cold temperature with an accumulated temperature of below 1,900°C (Cao et al., 2014). Lower soil surface temperatures and high humidity also promote the formation of plant tissue, leading to an increase in spore density and the accumulation of pathogenic Fusarium fungi, which in turn affects soybean disease (Yan and Nelson, 2022). Additionally, environmental factors influence both the pathogenic mechanism of the pathogen and the resistance pathways of the plant (Cheng et al., 2019).
This study provides the first systematic reports on the distribution, prevalence, and aggressiveness of Fusarium root rot soybean disease among the six ATZs in Heilongjiang province. Our results confirmed that the diversity of Fusarium spp. varies among different ATZs, but F. oxysporum and F. solani are the most frequently isolated species. Using a combination of morphological characterization and molecular analysis, we identified nine Fusarium species associated with soybean root rot, all of which can infect soybean plants with varying severity and affect plant biomass. F. oxysporum was found to be the most aggressive in terms of disease severity and impact on plant biomass, while F. brachygibbosum showed the second most inhibitory effect on root length after F. oxysporum.
RAPD analysis allows a better understanding of the pathogen’s virulence, evolution, and population genetic variation. Our results revealed relatively small genetic differentiation within populations, indicating significant genetic homogeneity within the six ATZ populations. This homogeneity is also reflected in the similar aggressiveness of the same species from different ATZs. These findings are consistent with previous studies, which demonstrated that F. oxysporum isolates exhibited less polymorphism and a high degree of genetic similarity among them (Nasr Esfahani, 2018, Brizuela et al., 2021). The low genetic diversity within F. oxysporum populations may be attributed to the narrow region in this study or the strictly asexual reproduction of F. oxysporum and limited gene flow between populations (Sillo, 2022).
In addition to temperature, numerous other environmental factors can influence the interactions between plants and microbes and disease development (Liu et al., 2020). Our study demonstrated that neither warmer nor colder conditions increased the diversity or richness of Fusarium spp., as reflected in ATZ1 or ATZ6. Instead, moderate temperatures appeared to favor the growth of Fusarium spp., as reflected in ATZ2. Many studies have reported that, under both low and high temperature regimes, Fusarium spp. exhibit low average mycelial growth rates (Saleh et al., 2021; Scott et al., 2010). Different Fusarium spp. exhibit diverse responses to temperature, indicating the complex genetic and evolutionary mechanisms underlying Fusarium species. Further research is necessary to explore the relationship between temperature, the pathogen, and the host, as this information will play a crucial role in managing diseases more effectively.
In conclusion, our study confirms that F. oxysporum is the dominant species causing soybean root rot in Heilongjiang province, distributed across six ATZs. For hemibiotrophic pathogens such as F. oxysporum and F. solani, cultivating resistant soybean cultivars would be the best control strategy. In our study, we tested 21 soybean cultivars for their resistance to the dominant and most aggressive species, F. oxysporum and F. solani. The same cultivar may exhibit a different response to F. oxysporum and F. solani infections, suggesting that different infection mechanisms may exist for different Fusarium spp. DN51 demonstrated resistance to both F. oxysporum and F. solani infections, making it a potential resistance material for future breeding programs or further studies on Fusarium-soybean interactions.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Supplement1
Supplement1 Print
Print






